Course Rationale
The field of watershed science, and specifically the study of streams, is inherently multidisciplinary, involving a broad array of physical, biological and social sciences. Traditional hierarchical undergraduate and graduate education programs to train students in the fields that support watershed science typically emphasize in-depth study within a specific discipline. This focused education is vital to producing professionals with useful technical and analytical skills. However, most students who pursue careers in watershed and stream analysis rarely work solely within their discipline. Rather, their work is inevitably integrated with that of other professionals addressing related issues with different skill sets. The ability to work closely and collaboratively with professionals from different backgrounds is fundamental to success in the field of watershed science.
This course introduced advanced undergraduate and graduate students to multidisciplinary collaborative watershed and stream analysis through combined laboratory and field study. Students from diverse backgrounds worked in cooperative research teams to collect and analyze field data from the Skeena River watershed (British Columbia), one of the largest un-dammed rivers in North America (Dynesius and Nilsson 1994). These teams used field collected data to analyze geomorphic processes and test ecological hypotheses related to the patterns of aquatic macroinvertebrate and salmonid diversity in tributaries to the Skeena River.
Executive Summary
Surveys of the downstream portions of tributaries to the Skeena River focused on the geomorphological and biological importance of stream confluences. Our key observations are listed below:
-
Tributary Inputs: Tributaries have a significant influence on the mainstem through input of sediment, coarse material, and large woody debris. These inputs directly affected the local geomorphology of not only each tributary but, in many cases, that of the mainstem channel. These inputs also influenced the spatial patterns of macroinvertebrates and salmonids. The upstream character of each tributary had a large impact on the character of the confluence.
-
Tributary Fans: A fan-shape deposit of river bed material forms at the tributary’s confluence with the mainstem. This tributary fan was delineated at the mainstem’s high water mark where sediment size and channel character (confined versus unconfined) changed abruptly. Fan dimensions were proportional to the mainstem channel width and stage-discharge relationships.
-
Stream Morphology Drivers: Habitat complexity in tributary streams was related to the presumed production and long residency time of large woody debris. Additionally, bedrock features created habitat complexity by altering the channel morphology.
-
Diversity Patterns: Macroinvertebrate and juvenile salmonid diversity were roughly equivalent in main tributary channels to those in the downstream tributary fans, although habitat complexity was greater upstream.
-
Macroinvertebrates: Larval heptageniid mayflies were the most abundant insects in the tributaries. The EPT index, often used as a measure of disturbance and stream suitability for salmonid rearing, was lowest in streams with high levels of disturbance due to the presence of beaver dams and large woody debris.
-
Salmonid Fish: The number of species observed or collected in or near tributaries increased as we moved down river. Young of year salmonids, especially coho salmon, steelhead, and cutthroat trout, were the most common fish in tributaries and fans, while larger fish were found more often in the main river at the confluences. Bull trout in particular exploited food resources at confluence zones. Juvenile salmonids seemed more abundant in the main river than in the tributaries.
Introduction
Recent investigations in geomorphology and riverine ecology have underscored the important physical and biotic interactions at stream confluences (Rice et al 2001, Benda et al 2004). From headwaters to confluence, biotas are usually thought to vary continuously in response to the constantly changing physical attributes (Vannote et al 1980). However, tributary inputs in the form of sediment, large woody debris (LWD), and biotic inputs such as detritus, invertebrates and juvenile fishes, generate punctuated interruptions to the hypothesized main stem continuum. Furthermore, mainstem dynamics have the potential to locally affect tributary physical structure and subsequent fish and macroinvertebrate distributions. These zones at confluences have a distinct physical as well as biotic character and present a well-defined unit of study. Although the effects of tributaries on the main stem have been hypothesized (i.e. link discontinuity concept of Rice and colleagues (2001) and network dynamics hypothesis of Benda and colleagues (2004)) only a few studies have quantitatively observed and described their character. With this report we aim to summarize our observations and measured attributes that describe the physical processes and biotic responses at sites where small tributaries flow into the larger Skeena River (river confluences). Our goal was to develop a first order understanding of the importance of these river confluences to aquatic biota, mainly salmonid fishes and aquatic macroinvertebrates.
The network dynamics hypothesis (NDH) argues that the physical characteristics at stream confluences create greater spatial and temporal habitat heterogeneity, which in turn create biological hotspots (Benda et al 2004). Tributaries alter both channel morphology by increasing sediment and LWD inputs by increasing the magnitude and frequency of disturbances. For example, habitat complexity can be increased by tributary inputs into the main channel. Sediment inputs into the mainstream create a backwater effect upstream from the confluence and correspondingly create a steep slope section of water just downstream. This change is gradient produces a tributary-forced pool and cascade zone surrounding the confluence. In addition, disturbance at confluences are more frequent due to asynchronous flooding between streams (i.e. localized flooding). A varied disturbance regime on the tributary as well as on the mainstem has the potential to increase aquatic diversity (Sousa 1979). Moreover the character of the confluence, including both watershed (i.e. relative basin sizes) and local factors (bedrock, angle of entrance, and location on the main stem meander bend) affect the morphology of the confluence.
A distinctive example of how tributaries influence the morphology of the mainstem is the presence of tributary fans. Tributary fans occur when the stream power of the tributary is large enough relative to the mainstem so that it transports more material than the mainstem removes locally. This causes a build up of material at the confluence. The dimensions of tributary fans vary depending on the character of the joining streams and their associated watersheds. Little has been quantitatively describe about these geomorphic features.
Tributary fans have a distinct set of physical parameters that create habitat for both aquatic macroinvertebrates and salmonids. The zone between low- and high-water is called the aquatic-terrestrial transition zone (ATTZ) (Junk et al 1989). It has been argued to be important for terrestrial resource input into the stream system and for rearing habitat for various fish species (Junk et al 1989). At confluences, short reaches exist that are perennially inundated by either mainstem or tributary waters; at lower flows, tributary waters flow over the confluence fan and at higher flows the main stem inundates the area. We are terming this perennially wetted area the mainstream-tributary transition zone (MTTZ). The aquatic biota may respond to the microhabitats created by the unique physical attributes of the MTTZ on the confluence fans. Little has been quantitatively observed in these areas regarding the spatial structure of aquatic organisms.
According to the NDH, network nodes, or confluences, offer the highest spatial heterogeneity and disturbance frequency, and therefore we should see a higher diversity of aquatic organisms. When comparing patterns of biological diversity, it is unclear whether the adjacent upstream portion of the tributary or the wetted confluence fan would have a higher diversity. On one hand, one could argue that tributary fans should have a higher diversity because they are connected to the main stem channel and thus have more connectivity to source populations. Conversely, tributary fans have a more frequent and higher magnitude disturbance regime than reaches not affected by main stem flooding. Therefore, habitat complexity should be higher in the upper reaches as residence time of LWD is longer. In addition, the presence of deeper pool riffle sequences in the confined section upstream of the fan offer better salmonid habitat than do several smaller channels braided over the confluence fan (i.e. MTTZ).
We hypothesize that both juvenile salmonid and aquatic macroinvertebrates species richness is higher in the upstream portions of the tributaries compared to the aquatic habitat on the tributary fan. Diversity in the tributary should be higher due to habitat complexity provided by large woody debris and decreased disturbance. We tested this hypothesis by surveying various tributaries to the Skeena River in British Columbia, Canada where tributary fans are abundant along the mainstem. In addition, the study describes the geomorphology of tributary fans, examines the physical processes that create them, and describes the spatial structure of salmonid fishes and aquatic macroinvertebrates at stream confluences.
Study Area
The Skeena River watershed is about 54,000 km2 and extends 570 kilometers from central British Columbia to the western Pacific coast at the city of Prince Rupert. The Skeena River is the second largest river in British Columbia and is one of the longest un-dammed rivers in the world (Dynesius and Nilsson 1994). Glacial advance and retreat, driven by climate cycles, has sculpted the valleys and canyons of British Columbia and directly affected rates of erosion of the mountains. The majority of the Skeena River lies within the intermontane belt. This belt is made of uplifted marine sedimentary rocks that are more conducive to weathering than either igneous or metamorphic rocks. The river channel often encounters folds of erosion- resistant sandstone and mudstone, which causes the river to narrow into deep gorges. In the downstream reaches, the Skeena River enters into the coastal belt, which consists of erosion-resistant granitic rocks.
The Skeena River is a diverse watershed that receives water from glacial ablation, snowmelt, and precipitation as rain. Hydrologic inputs are dominated by snowmelt and glacial ablation from mid spring to late summer where flows are historically as high as 9000 cms and average about 3500 cms. A second flow peak occurs between October and December when precipitation falls as rain because it is not cold enough for snow. These autumn flows are historically as high as 5500 cms and average about 2500 cms. The dominant vegetation in the watershed is Sitka Spruce, Subalpine Fir, Quaking Aspen and Black Cottonwood forests. Between December and May river flows are at a minimum.
The Skeena River watershed is famous for its anadromous fish runs of up to 5 million salmon and steelhead each year. The river currently supports extensive commercial and sport fisheries and has played a major role in the livelihood of Native Peoples living within the Skeena watershed. Tribes within the Skeena watershed include the Northern Tsimshian, the Southern Tsimshian, the Gitskan, and the Nisga'a. Although large tracts of the watershed are relatively unaffected by anthropogenic influences, logging, mining and farming activity are deemed to be a significant threat to steelhead and salmon productivity (FOC 2002).
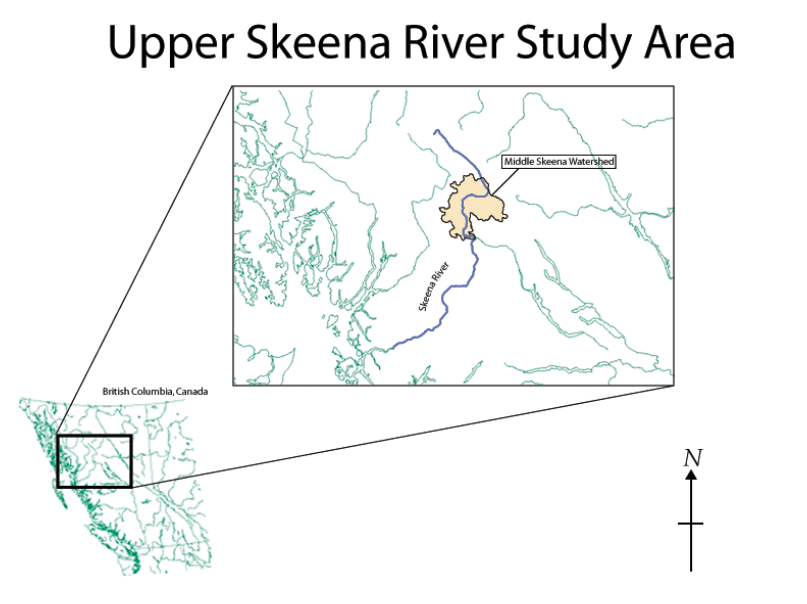
Our study area was located within the Middle Skeena Watershed downstream from Mosque Creek (Figure 1). While we observed the river from Mosque Creek to Kispiox, our survey efforts were focused in the upper reaches between the Squingula and Babine Rivers.
Methods
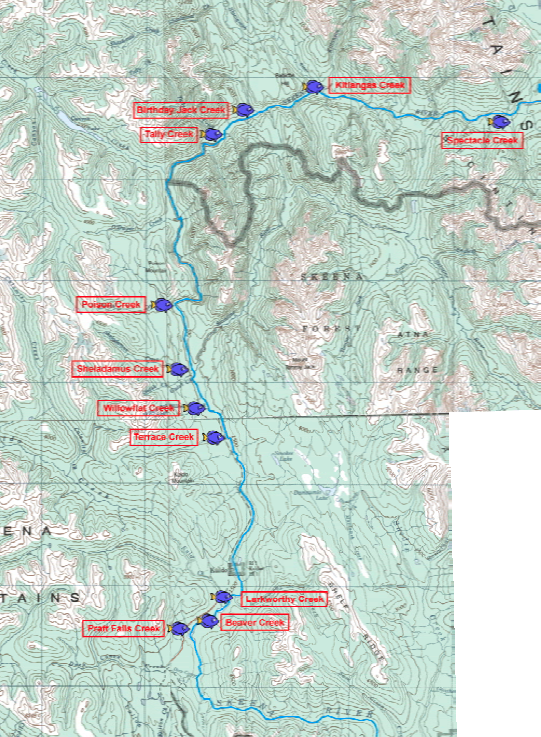
For the field study portion of the class, students were divided into two interdisciplinary teams of six people. Each team consisted of students with an assigned emphasis in one of the following fields: geomorphology, hydrology, fisheries or aquatic ecology. Eleven streams tributary to the Skeena River within the Middle Skeena watershed were studied in detail in order to assess differences between the tributary fan and the confined tributary reaches upstream (Figure 2). One team characterized the confluence characteristics and the corresponding tributary fan, while the other team characterized a confined reach upstream of any confluence effects. Each team completed a reconnaissance of their selected stream site to assess the overall character of the stream habitat and the diversity of the channel morphology in order to determine a representative reach to characterize. Areas within each study segment were classified into geomorphic types based on the classification of Montgomery and Buffington (1997). Specifically, stream reaches within each segment were classified as one of five morphologic types: bedrock, cascade, step-pool, plane-bed, or riffle-pool. Geomorphic, macroinvertebrate, and fish surveys were then conducted in order to more fully determine quality of salmon spawning habitat. Details on the specific survey methods used to characterize each sample reach are described below.
The methods utilized in this course combined techniques primarily from the U.S. Geological Survey's National Water Quality Assessment Program (USGS NAWQA) (Fitzpatrick et al 1998), the U.S. Forest Service's Field Guide to Stream Channel Reference Sites (USFS) (Harrelson et al 1994) and the Environmental Protection Agency's Rapid Bioassessment Protocol (EPA RBP) (Barbour et al 1999) with the specific intent of evaluating several factors that may limit anadromous fish spawning and rearing success. Those factors included stream habitat availability (channel structure and flow conditions), water quality, and relative abundance of food sources (macroinvertebrates). Although many other limiting factors exist for anadromous fish spawning and rearing success, time limitations associated with this class precluded further analysis.
Reach Characterization
The reach characterization of each tributary studied was a qualitative investigation of the geomorphic and biotic character of the reach. Based loosely on the 'reach' form used in the USGS NAWQA protocols, a 'reach characterization' data sheet was used to document the physical conditions and characteristics of the sample reach. These general characteristics were chosen as factors that may directly or indirectly affect salmon spawning and rearing habitat. Recent and current weather conditions were noted on this form along with stream morphology, sediment sources, and anthropogenic impacts. Presence or absence of large woody debris, aquatic vegetation, and dominant vegetation within the riparian corridor play a major role in the heterogeneity of salmon and invertebrate habitat and were noted in the reach characterization. Water quality parameters and stream type (glacial, organic rich, or clear) were also noted. Lastly, stream flow was estimated using a velocity area method and the float stick method (Harrelson et al 1994).
For each reach a detailed map of the geomorphology, flow morphology, and vegetation was drawn. The reach sketch included the major facies (rock size types), the locations of the cross sections, and the placement of invertebrate kick-net samples. It also included any major woody debris and showed how debris and bedrock affect channel morphology and, presumably, salmon habitat.
Geomorphic Surveys
The geomorphic characteristics of each sample reach were evaluated using the USFS protocols described by Harrelson et al (1994). The geomorphic surveys were conducted to capture the geometry of the tributary channel and of the surrounding geomorphic features such as terraces and floodplains. There were two major components to the geomorphic survey, the cross section and the longitudinal profile. These surveys were recorded on the ‘channel morphology data sheet’. Two cross sections were chosen to represent the channel geometry at the dominant geomorphic types. For example, if the stream reach was a riffle-pool system then one cross section was taken across a representative riffle and one was taken across a representative pool. The longitudinal profile was surveyed along the thalweg and depicted localized change in channel bed slope as well as the overall slope of the reach.
The ‘sediment characterization data sheets’ were used to quantify the facies types that were delineated in the reach sketch. The facies types were collections of various sediment sizes that were found in the reach. For example, a reach may have contained three collections of grain size: a sandbar, a bar of well-sorted small cobbles, and a bar of poorly-sorted medium and large sized cobbles. Three facies types would be delineated in the reach sketch and each facies type would be quantified on a sediment characterization form. Each facies type was quantified using the Wolman pebble count procedure (Wolman 1954), and standard grain size variables (D16, D50 and D84) were determined from a cumulative percentage curve.
Macroinvertebrate Surveys
Benthic macroinvertebrates were sampled following the EPA RBP protocols for a single habitat (Ch.7, Barbour et al 1999). Samples were focused in riffles or runs where macroinvertebrate abundance and diversity is usually highest due to the prevalence of coarse substrate material. Two or three kicknet samples were taken per reach using a D-frame kicknet. A single kick sample consisted of disturbing the sediment to the depth of the largest particle in front of a kick-net for 90 seconds at each sample site. Samples were sorted and identified onsite, and any unknown individuals were stored in ethanol for later identification to family using standard keys. Data was recorded on a 'Macroinvertebrate Survey Datasheet' that included information collected at each kick sample site (such as local velocity, depth and temperature at the kick site) as well as the taxa identification data collected during the post-processing. Sample locations were also delineated on the reach sketch form for that sample reach.
EPT taxa and EPT index were used as indicators of reach disturbance and likely suitability for rearing juvenile salmonids. The EPT index is calculated by summing counts of total combined individuals in the orders Ephemeroptora, Plecoptera, and Trichoptera and then dividing by the total number of individuals of all invertebrates in the sample. The EPT taxa count is simply the number of families of Ephemeroptera, Plecoptera, and Trichoptera observed in the kicks.
Fish Surveys
Relative fish abundance and diversity in each sample reach was evaluated from a snorkel survey of the entire reach length. One snorkler would survey the length of the reach calling out observed species type, number of individuals, approximate size (in fork length) and associated habitat type to a data recorder on the stream bank. Data were recorded on the 'Fish Survey Datasheet'. In an effort to augment the snorkeling surveys, seine nets were pulled through pools, riffles, and backwater areas. Captured fish (predominantly juveniles) were identified to species, and fork length was recorded in field notebooks.
Key Findings
Geomorphology
Tributary Fans
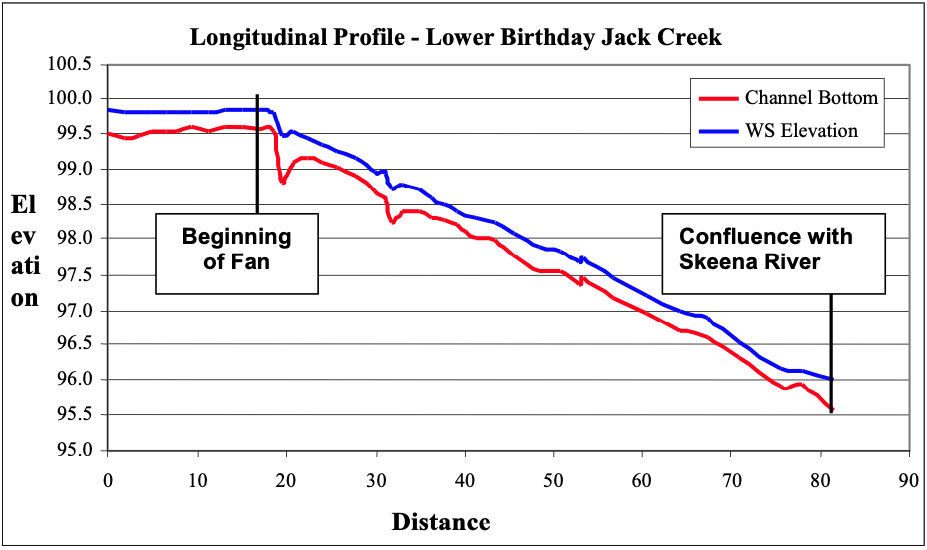
On the upper reaches of the Skeena River, tributary fans are a signature feature of many of the tributaries. Figure 3 shows an idealized tributary fan in both planform and longitudinal view. As the tributary flows into the mainstem of the river, the channel becomes unconfined with an abrupt increase in slope. This transition marks the head of the tributary fan and corresponds with the high water mark of the mainstem. For this project, the high water mark was determined visually by noting the edge of the riparian (namely cottonwood and alder) and the aquatic vegetation (periphyton and algae). The correlation between the break in slope and the edge of vegetation shows that the base elevation of the tributary is predominately controlled by the high-stage of the mainstem. That is, the tributary base level corresponds to the effective discharge of the tributary and corresponding high flows on the main stem. As water recedes, the tributary alluvial deposits not yet removed by the mainstem flows lay exposed on the channel bank. They take a fan-like form with some additional sediment deposited by the mainstem on the leeward side. The fact that a relationship between the tributary and the mainstem exists shows that the slope and height of the fan are directly related to the channel width and stage-discharge relationship of the main stem. The abrupt change in slope is evident in the longitudinal profiles of each creek shown in Appendices 1-11, but most notably on the profile of Birthday Jack Creek (Figure 4).
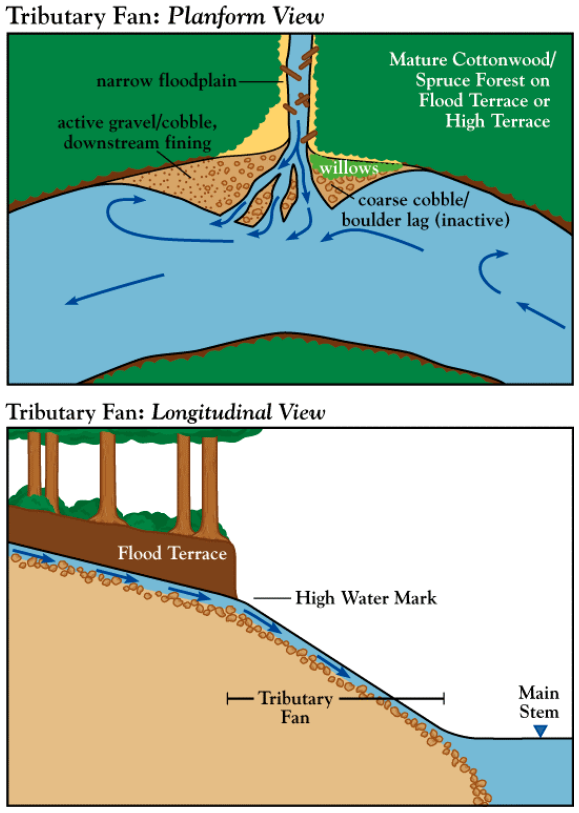
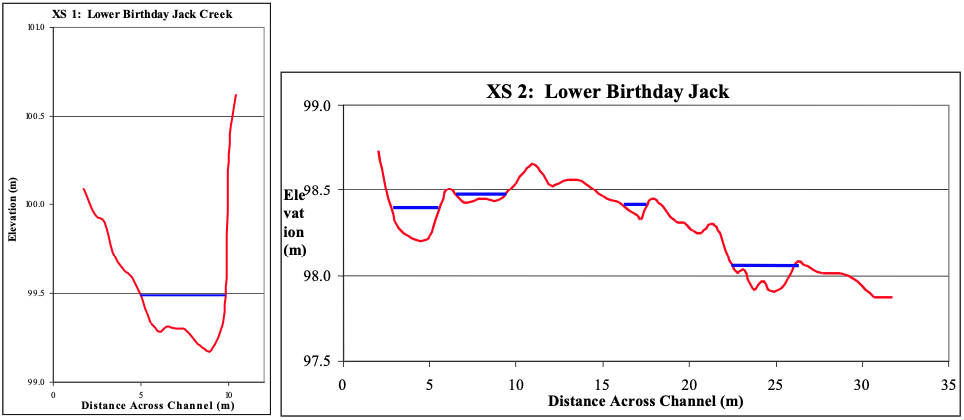
In cross-sectional view, the confined reach of the tributary has a normal channel form (See Figure 5). A cross-section view across the foot of the fan reveals in general a hump form with the smaller braided channels. A good example on this form can be found in the lower section of Birthday Jack Creek in Appendix 3 (Figure 5). The humped shape is caused by sediment input from the tributary and subsequent deposition and scour from the main stem. Sediment on the tributary fan becomes organized due mainstem flow. On the tributaries with larger fans, imbrication of the surface particles was evident. Larger sized facies occurred at the upstream portion of the fan with pebble to sand sized material deposited by the main stem on the downstream portion (relative to the Skeena River) of the fan. This is shown in Figure 3 where smaller sediments are deposited on the lee side of the tributary fan.
Tributary fans with the above described features have unique hydrologic characteristics. During high flow the tributary fan is inundated by mainstem water and during low flow by tributary water. On the Skeena River, in particular, tributary waters can have a very dissimilar character. Many of the tributaries we observed were clear, cold water streams with low discharges relative to the main stem. Skeena River water is cloudy and turbid due to glacial inputs and high sediment loads from upstream tributaries. Two other types of tributary waters were observed, wetland and glacial sourced waters. Each type has a unique impact on the local ecology (See macroinvertebrate and salmonid fish sections for more detail).
The transition zone that occurs at tributary confluences with its distinct geomorphology and hydrology creates an environmental ecotone to which fish and aquatic macroinvertebrates most likely respond. In the main stream, opportunistic predatory fish such as bull trout (Salvelinus confluentus) will exploit fish, invertebrates, and other prey associated with the tributaries and their fans. The rivers below fans are attractive to feeding salmonids presumably because the tributary delivers food organisms to the river and because the fish can use the more turbid water of the river as cover to exploit small fish and other organisms in the clearer tributary water (Moyle 2002 and P Moyle, pers. obs.). On the tributary side of the confluence little information exists prior to this study. For example, tributary fans at low flows have the potential to block entrance of predatory fish into tributaries or may hinder the movement of fish up for spawning. The role of tributary fans in regulating upstream populations of fish is clearly a subject that needs further exploration.
Stream Morphology Drivers
Observations and qualitative data show that large woody debris has a tremendous impact on tributary stream morphology and thus upon salmon habitat. In almost every instance where large woody debris was plentiful, stream morphology tended towards a step-pool or riffle-pool character. When large woody debris falls into a tributary it constricts the channel forcing flow velocity to increase around or over the debris. When velocities increase around the debris, scour occurs along the leading edge of the debris and a depositional eddy forms on the downstream side of the debris. When water flows over the debris, a depositional bar forms on the upstream side of the debris due a backwater effect and scour occurs on the downstream side of the debris- formed step. In both cases the widening of the channel downstream of the debris leads to decreased velocities, deposition of sediment and in some cases pool formation. Over half of the tributary reaches surveyed had stream morphologies significantly impacted by woody debris resulting in forced step-pool formations.
A comparison of the reach sketches reveals that pool formation is much more common on tributaries with extensive woody debris. Specifically, velocities are lower and depositional features are common on the downstream edge of the woody debris. A good example of this process is shown at the upstream end of the survey reach on Birthday Jack Creek (Appendix 3). The reach sketch map and thalweg profile show that water depth increases on the downstream
side of woody debris. The woody debris dam and subsequent downstream pool at the top of the reach is the most pronounced example. In stark contrast to the upper confined tributary reaches, the tributary fans were devoid of woody debris resulting in channel morphologies dominated by riffles and cascades with minimal to no pool formation.
Bedrock control was another significant driver of stream morphology. Bedrock outcrops of erosion-resistant rocks, such as sandstone, would often constrict the channel width increasing velocity and forming bedrock steps and chutes. In a similar fashion to that of woody debris, forced step-pools and riffle pool sequences would form downstream of the bedrock outcrop. One example of this was on the upper reach of Poison Creek (Appendix 5). Both the bottom and top of the reach contained bedrock outcrops that formed steps and associated scour pools immediately downstream. Further downstream of the immediate influence of the bedrock constriction, the channel generally widens resulting in a shallow riffle or run (see thalweg profile in Appendix 5).
Sheladamus Creek (Appendix 6) was a unique tributary that exhibited strong controls from both large woody debris and bedrock. The creek passed through a canyon with steep bedrock cliffs on either side and contained extensive large woody debris from fallen snags. Many of the fallen snags had created forced step-pools; however, lee deposits and pool infilling were minimal suggesting that high flows from snowmelt may cause extensive scour and periodically entrain the woody debris. As a result, woody debris may control stream morphology during low flow, while bedrock influences stream morphology at high flows over longer time scales.
With respect to salmon habitat, woody debris and bedrock constriction are key drivers in creating deep shady pools for cover and forcing deposition of gravels necessary for spawning. These features also provide excellent rearing habitat for anadromous and non-anadromous species alike. The tributaries where large woody debris or bedrock outcrops forced pool-related morphologies were not only more diverse in habitat type, but exhibited greater species abundance and richness.
Invertebrates
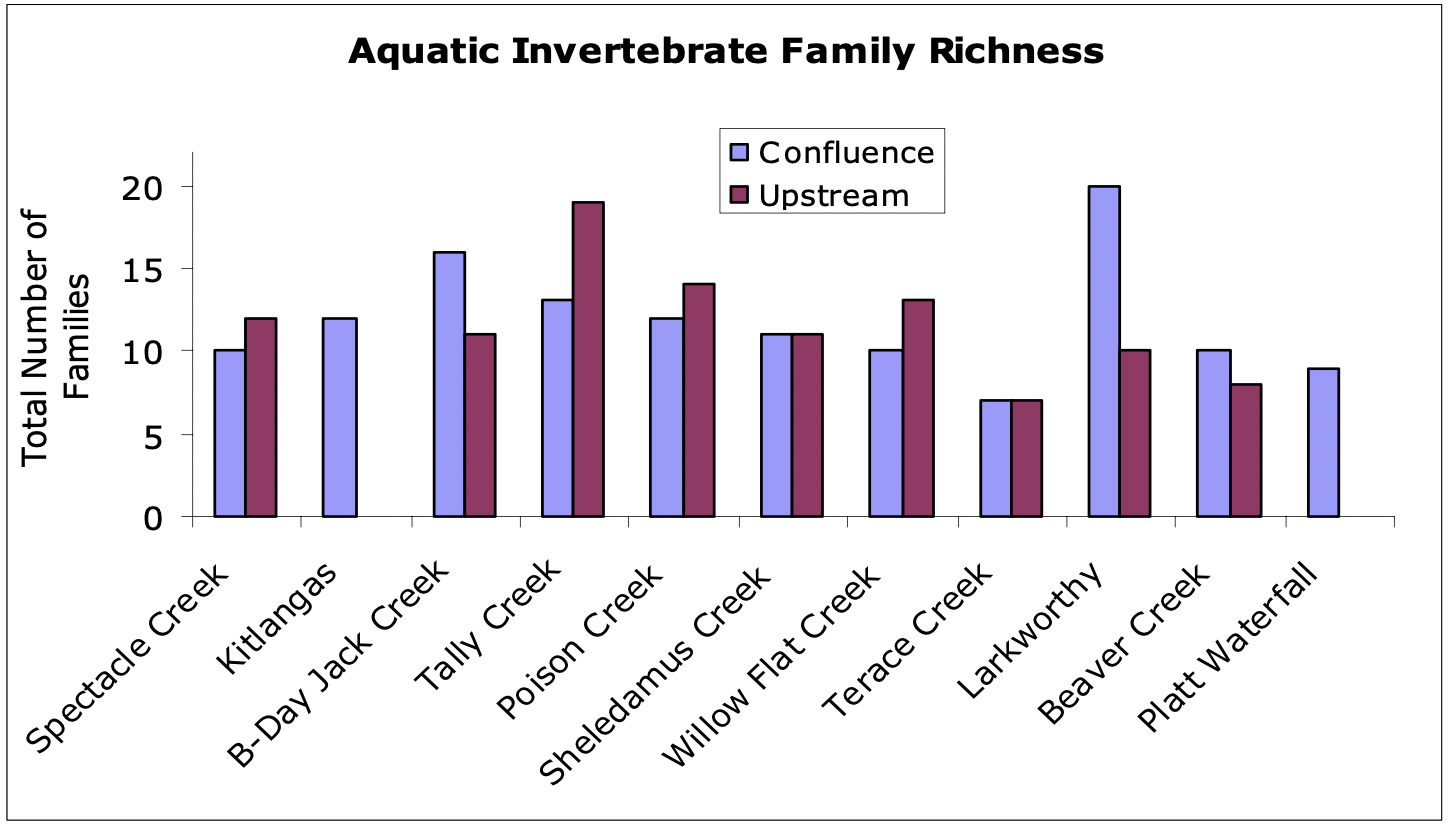
Between seven and twenty families were identified in each reach from the 11 tributaries. In total, seven ephemeropteran (mayflies) families, four plecopteran (stoneflies) families, four trichopteran (caddisflies) families, and six different dipteran (true flies) families were identified. In addition, mites (hydracarinids), platyhelmenthids (flatworms), mollusks (snails), coleopterans (beetles) and annelids were occasionally observed. Due to the field identification methods, some taxa were likely lumped or overlooked, so family counts may be conservative.
On average, 44 invertebrates were collected at each kick (about 0.5m2 in area) throughout the surveys. A significant exception was Beaver Creek. This slow moving creek with pebble and small cobble wetted facies had low invertebrate counts dominated by dipterans, which tend to thrive in more disturbed areas and in areas with finer substrate. Tributary modification by beavers and potential water quality differences due to the large wetland above a beaver dam complex were possibly contributors to the low invertebrate abundance and diversity at this site.
Mayflies
Heptageniids, which are primarily scrapers and gathering collectors that live in fast- moving water, were abundant in every stream except Beaver Creek. Relatively large siphlonurid mayflies were found in the greatest abundance in Willow Flat Creek and Sheladamus Creek; individuals from this family are collector-gatherers and predators. Their large size indicates stability of flow levels in these creeks to allow time for their longer life cycles. Adult mountain whitefish and bull trout captured in Sheladamus Creek were feeding heavily on the sipholonurid mayflies.
True flies
Chironomids, which are gathering collectors, were especially abundant in the confluence reaches of Spectacle, Tally, and Larkworthy Creeks, while they were not as common in the upstream reaches of these rivers.
Caddisflies
Case building trichopterans were observed in small numbers in all but two confluence reaches. The greatest abundance and diversity of case-building trichopterans was in the shallow, large cobble regions of Pratt Falls Creek. The predatory free-living Rhyacophilidae were found in all streams but Terrace, Beaver and Pratt Falls Creeks.
Stoneflies
Plecopterans are adapted to well oxygenated streams and rivers as nymphs have fixed gills that can only extract oxygen in moving water. The largest variety of stonefly families (5 or more) was found in riffles from Tally and Larkworthy Creeks at water velocities of 0.7 and 1.3 m/s respectively.
Families
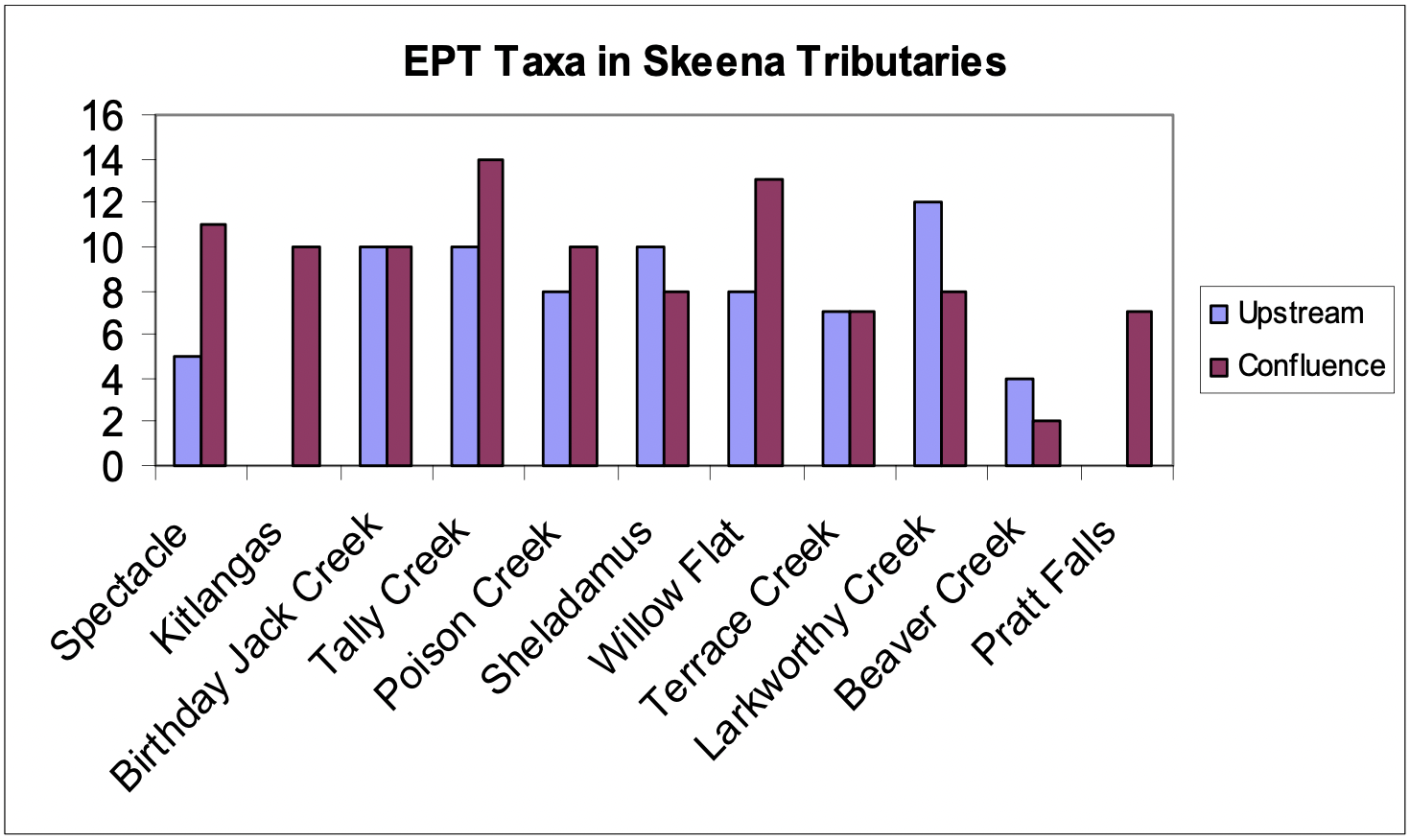
We found the largest number of families in Tally Creek and Larkworthy Creek. These large tributaries had both an abundance of large woody debris within the channel and broad low gradient alluvial fans. These tributaries with large watersheds, higher flows and thus greater transport capacity had more power to adjust the slope of their confluence with the Skeena River than the smaller creeks we surveyed.
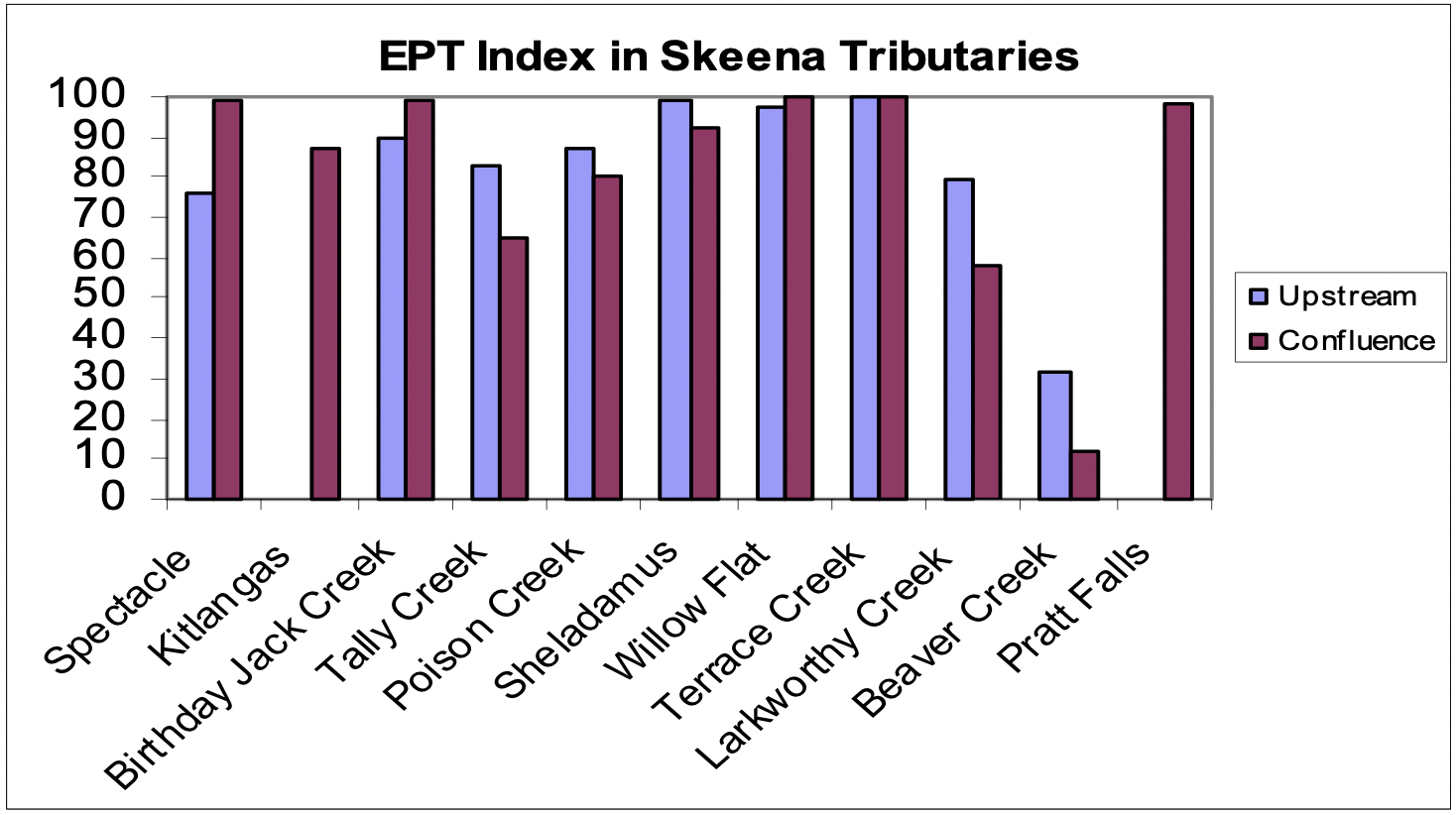
The EPT (Ephemeroptera-Plecoptera-Trichoptera) index, which is commonly used as a measure of water quality, disturbance, and salmonid food availability, ranged from 12 in the slow backwater area of Beaver Creek to 100 in Willow Flat Creek and Terrace Creek. The EPT index values were similar between confluence and upstream reaches for most of the tributaries (Figure 6). The high level of disturbance due to the beaver dam buiding, slow water velocities and small cobble/gravel substrate likely all contribute to the poor EPT index at both Beaver Creek reaches. In comparing streams, the EPT index did not follow a particular trend. With the exception of Larkworthy, Sheladamus, and Beaver Creeks, the number of EPT taxa in confluence reaches was greater than or equal to that in the upstream reach in 6 creeks, while the number of EPT taxa was lower in the confluence than in the upstream reach in three creeks (Larkworthy, Sheladamus, and Beaver Creeks) (Figure 7).
Additional infomation that should have been collected in our invertebrate survey is the local scale facies type at the exact location where invertebrate samples were taken. We noted large cobble substrate often had a higher number of families and larger individuals than did small cobble or gravel. Limiting our sampling sites to riffles in all reaches would have also be helpful to more accurately compare creeks and reaches.
At the outset of this study, we hypothesized that greater invertebrate diversity would be found in upstream habitats due to greater habitat complexity and a less severe disturbance regime. We found, however, that family richness did not follow a consistent pattern between confluences and upstream reaches in the tributaries (Figure 8). We conducted a canonical correlation analysis (CCA) to determine whether some families were more commonly found in either confluences or upstream reaches and whether water depth and velocity are particularly important in determining the location of families. The CCA revealed no significant clustering of commonly observed families between confluence and upstream reaches. In general, upstream reaches were associated with lower velocities than confluence reaches. Rare species including snails, mites, annelids and simuliids tended to be found in the upstream reaches. Trichopteran and chironomids tended to be associated with the confluence more often than with upstream reaches. The eleven pupa observed were associated with higher depths and confluence reaches.
Salmonid Fish
Surveys of distribution and abundance of fishes of the Skeena River and its tributaries yielded only salmonids, including coho salmon (Oncorhynchus kisutch), sockeye salmon (O. nerka), Chinook salmon (O. tshawytscha), steelhead (O. mykiss), coastal cutthroat trout (O. clarki clarki), bull trout (Salvelinus confluentus), Dolly Varden (S. malma) and mountain whitefish (Prosopium williamsoni). Each species was observed in multiple age classes, and usually found in various habitat types within each sampled reach.
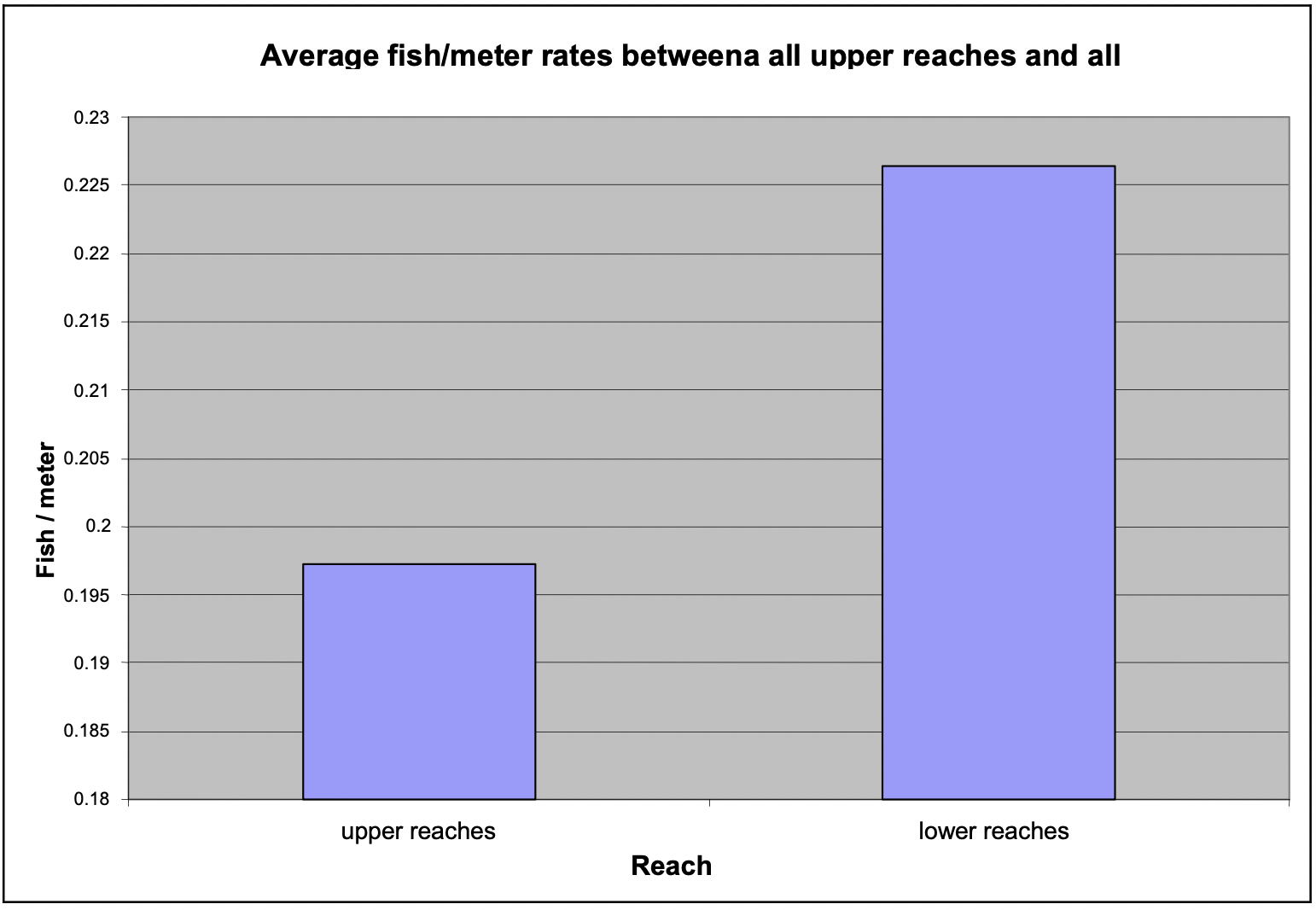
Fish densities (via fish/meter) between upper and lower reaches throughout our study were compared, and no significant differences were found (Figure 9). Fish found in the Skeena River at the confluence with the tributaries were omitted from calculations, because the fish were not truly in the tributaries being studied.
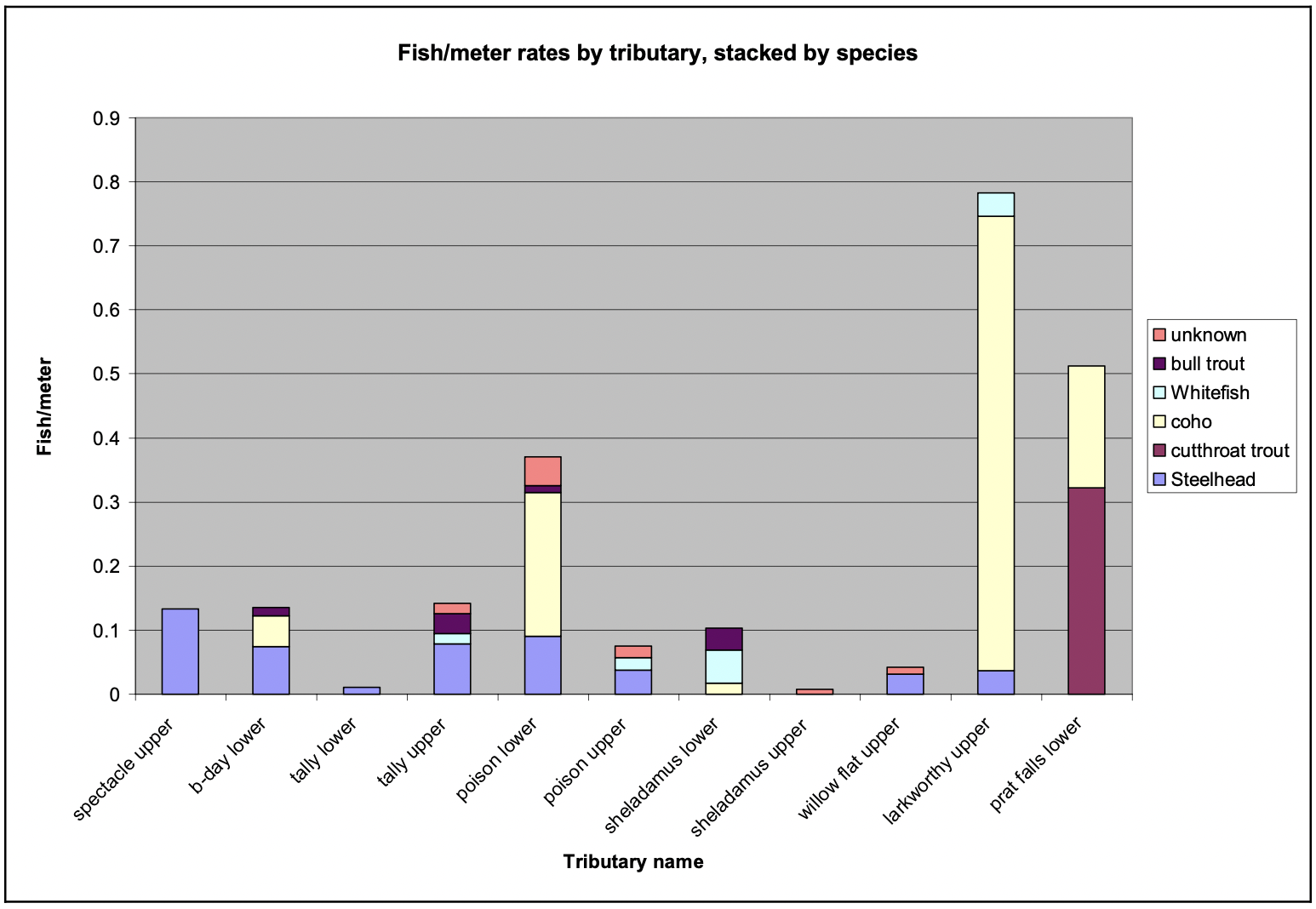
Coho Salmon
Coho were the most abundant species sampled, and were consistently observed in tributaries throughout the study, as well as in the mainstem. Most coho were observed in woody debris (64.5% of all coho found) and pools (27.9%), and small numbers were found in cascades (0.6%), riffles (0.6%), and edge habitat (1.2%). The remaining 5.2% of the coho were found in the mainstem of the Skeena River at the confluences of the tributaries. Of the ten tributaries that were surveyed, coho were observed in six. Furthermore, of the streams they were found to occupy, coho appear to be the most abundant of the species present as quantified by number of fish per meter (Figure 10).
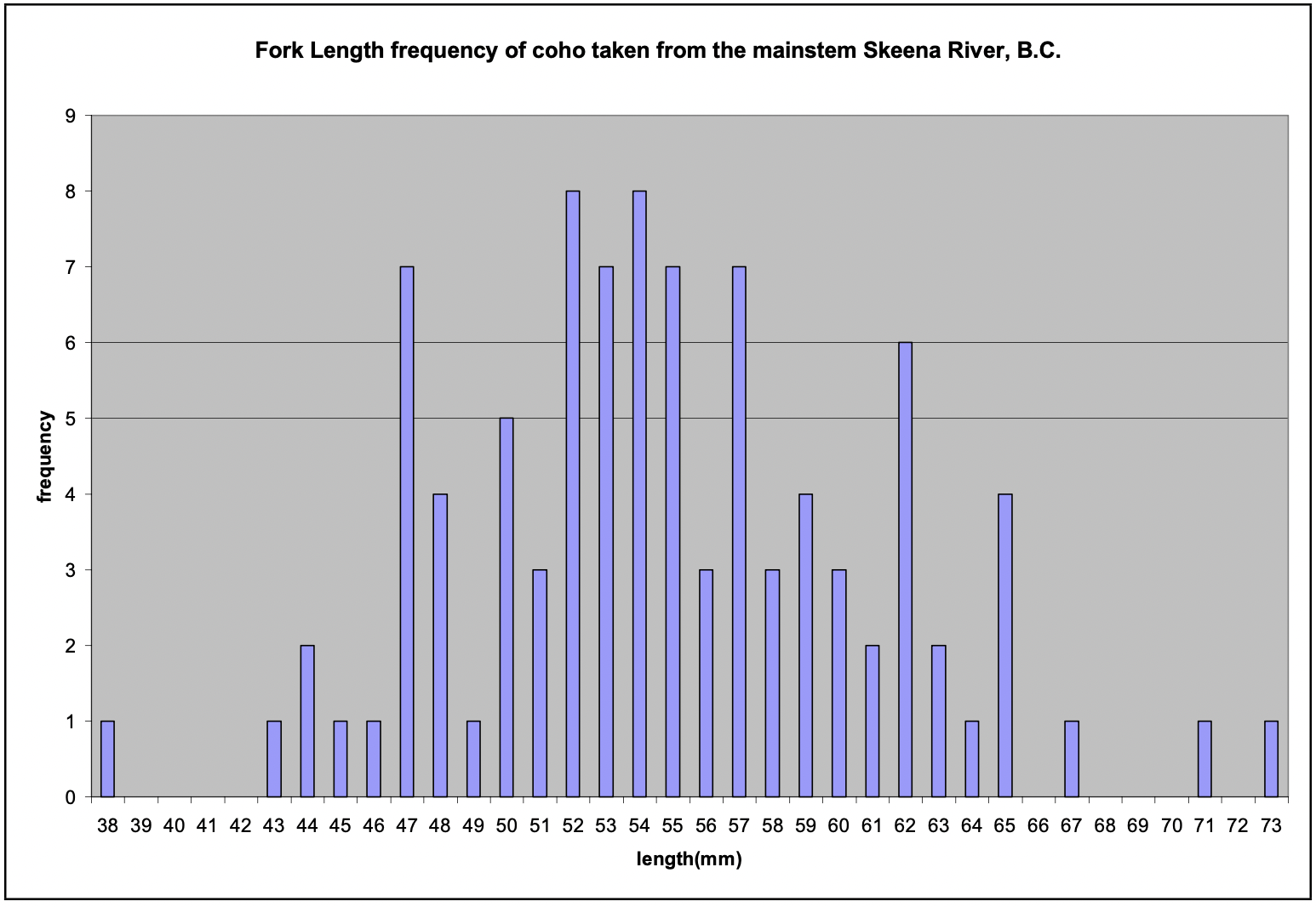
There were two length groupings of coho consistently observed in the tributaries. One group was less than 50 mm fork length (FL) (4.6% of total coho), and the second was between 50-100 mm (95.4%). Both size groups and habitat utilized were consistent throughout the study regardless of tributary or reach. Interestingly, for coho found in the mainstem of the Skeena River, there were no defined breaks in length classes, with very few fish found outside the 50- 100 mm size group (Figure 11).
Steelhead
Although not the most abundant species found, steelhead were observed in more tributaries (7 of the 10 surveyed) than any other species throughout the study. Snorkel surveys found steelhead to equally utilize pools (31%) and woody debris (31%) for habitat, and to a lesser extent use edge habitat (17.2%) and cascades (6.9%). Furthermore, 13.8% of the total steelhead were found at confluences between the tributaries and the Skeena River.
Steelhead were also found in two size/age classes; 55.2% were young of the year (<80 mm FL) and 44.8% were yearlings (>80 mm FL). A few (<1%) steelhead were observed in the size range 200-300 mm FL, indicating either a resident rainbow trout population or presence of two year old steelhead. Sampling bias and/or error may have limited the number of steelhead observed in this size class.
Mountain Whitefish
Mountain whitefish were found in 6 of the 10 surveyed tributaries. They were found to inhabit pools (31.3%), edge habitat (37.5%), woody debris (6.2%), and confluences between tributaries and the Skeena River (25.0%). There was no difference in habitat utilization in different tributaries among mountain whitefish.
Very few juvenile mountain whitefish were found in the tributaries, although they were present in the main river and in backwater areas. Only 18.8% of all whitefish were smaller than 100 mm FL, while the rest fell within two other size groupings. 43.8% of the whitefish were between 100 and 300 mm, and the remaining 37.5% were 300-400 mm FL.
Bull Trout
Bull trout were also found in 6 tributaries. While snorkeling in tributaries, 22.2% of the bull trout were found in pools, and 5.6% were found in riffles. The majority of bull trout were sampled by hook and line at confluences of the Skeena River and tributaries, where the remaining 72.2% were found.
Because the majority were sampled by hook and line, there is a bias for larger size classes in our data. Therefore, 72.2% were between 100 and 350 mm FL. However, 16.7% of the bull trout were smaller than 100 mm, and 11.1% were larger than 350 mm. For more information on bull trout during our study, refer to appendix 12.
Coastal cutthroat trout
Cutthroat trout were found in only 2 streams of the 10 surveyed, but were found in densities that were relatively high compared to other species found (cutthroat comprised 17.4% of the total fish found). Cutthroats were the only species found to consistently utilize riffle habitat, as 40.3% in our study were found there. However, 32.8% of the cutthroat also resided in edge habitat, 7.5% were in cascades, 4.5% were in pools, and 14.9% were found in confluences with the Skeena River and tributaries. We did not document any cutthroat until later in our study, possibly implying that they occupy lower reaches of the Skeena River than do the other fish we encountered. Almost all of the cutthroat (95.5%) were juveniles, under 50 mm FL, which typically occurred by themselves, in shallow water in deep cover. The remaining 4.5% of the cutthroat were adults, between 300-400 mm.
Sockeye Salmon
Sockeye were the least abundant fish found in our study, comprising only 4.2% of all fish sampled. Sockeye were found at only one tributary (Beaver Creek). 87.5% of the sockeye found were in riffles, while 12.5% were located in a slow-moving backwater area in the lower reach of the study site. Because our sample size is restricted, it is not possible to make any inferences regarding habitat preference. All sockeye sampled were juveniles under 50 mm FL. However, there were large pulses of spawning sockeye moving upstream in the mainstem of the Skeena River below the confluence of the Babine River. Five sockeye and a pink salmon (O. gorbuscha) >600 mm FL were collected by gill net by a local First Nations tribal member and given to us for culinary study. The study found that sockeye salmon were superior eating to pink salmon when roasted over coals, although the small sample size suggests that further study is needed.
Constraints
Studying the effect of tributaries on main stem dynamics is a more straight forward endeavor than reciprocal analysis on the tributary. Like all scientific inquiries, environmental complexity is easier to understand on paper than in the field. In addition, surveying the mainstream would have been too difficult to complete safely. Many times even the moderate- sized tributaries were too dangerous to actively survey, especially those with high flows from recent rains. These constraints aside, this course produced both valuable observations on river system process and patterns as well as the opportunity for students to absorb and experience scientific inquiry as it is practiced, in the field.
While our data is incomplete and very fragmentary in nature, it does suggest that further study of these confluences is warranted, especially because they suggest the Network Dynamic Hypothesis has limited applicability to the Skeena River system. Future studies should involve more effective fish sampling (probably through electrofishing), increased numbers of confluences studied, and studies further upstream from the influence of the main river. Such studies could have additional value in increasing knowledge of the least understood fishes of system such as bull trout, cutthroat trout, and mountain whitefish.
Appendix
Appendix 1 - Spectacle Creek
Appendix 2 - Kitlangus Creek
Appendix 3 - Birthday Jack Creek
Appendix 4 - Tally Creek
Appendix 5 - Poison Creek
Appendix 6 - Sheladamus Creek
Appendix 7 - Willow Flat Creek
Appendix 8 - Terrace Creek
Appendix 9 - Larkworthy Creek
Appendix 10 - Beaver Creek
Appendix 11 - Pratt Falls
Literature Cited
Barbour, M.T., J. Gerritsen, B.D. Snyder, and J.B. Stribling. 1999. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish, Second Edition. EPA 841-B-99-002. U.S. Environmental Protection Agency; Office of Water; Washington, D.C.
Benda, L. N., L. N. Poff, D. Miller, T. Dunne, G. Reeves, G. Pess, and M. Pollock. 2004. The network dynamics hypothesis: how channel networks structure riverine habitats. BioScience 54:413-427.
Dynesius, M., and C. Nilsson. 1994. Fragmentation and flow regulation for river systems in the Northern Third of the World. Science 266:753-762.
Fisheries and Oceans, Canada (FOC). 2002. Pacific Salmon. http://www.dfo-mpo.gc.ca/zone/underwater_sous- marin/salmon/salmon-saumon_e.htm. Date accessed: 5/26/04.
Harrelson , C.C., C.L. Rawlins, and J.P. Potyondy. 1994. Stream channel reference sites: An illustrated guide to field technique. U.S. Forest Service, Rocky Mountain Forest and Range Experiment Station. General Technical Report RM-245.
Junk, W. J., P. B. Bayley, and R. E. Sparks. 1989. The flood pulse concept in river-floodplain systems. in D. P. Dodge, editor. Proceeding of the International large river symposium (LARS). Department of Fisheries and Oceans, Honey Harbour, Ontario, Canada.
Montgomery, D.R and J.M. Buffington. 1997. Channel-reach morphology in mountain drainage basins. Geological Society of America Bulletin 109(5):596-611.
Moyle, P.B. 2002. Inland fishes of California. Berkeley, University of California Press.
Rice, S. P., M. T. Greenwood, and C. B. Joyce. 2001. Tributaries, sediment sources, and the longtudinal organisation of macroinvertebrate fauna along river systems. Canadian Journal of Aquatic Sciences 58:824- 840.
Sousa, W. P. 1979. Disturbance in marine intertidal boulder fields: the nonequilibrium maintenance of species diversity. Ecology 60:1225-1239.
Vannote, R. L., G. W. Minshall, K. W. Cummins, J. R. Sedell, and C. E. Cushing. 1980. The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences 37:130-137.
Wolman, M.G. 1954. A method of sampling coarse river bed material. American Geophysical Union 35:951-956.