There are 17 known species of bats found in Yosemite National Park (Sears 2011, Stock 2015). Of these 17 species, 5 are listed as special status due to documented population declines (Pierson 1998). These declines have mainly been attributed to anthropogenic impacts outside the park, but park conservation efforts have provided essential refuge habitat and contribute to thriving bat populations. Bats are known to forage on numerous types of insects (Naidoo 2012, McWilliams 2005, Clare 2011) in respect to location, seasonality and habitat conditions. Several species of bats, including the majority of the species found within and near Yosemite National Park, have foraging strategies that are closely associated with bodies of water (Seidman 2001). Riparian areas and adjacent forested areas not only provide critical foraging habitat for bats, but also for roosting and mating (Brigham 1997). The rich riparian vegetation and seasonal ponds attract a variety of insects which bats feed and forage on (Clare 2011, Seidman 2001). The persistence of bats can serve as an indicator of habitat and ecosystem health by their presence and abundance. Bats are considered an indicator species because they must consume 30-50% of their body weight each night in order to survive (Esbérard 2008). Thus, in order for healthy populations of bats to persist, their habitats must have the appropriate conditions for roosting, open spaces for foraging, and plentiful insects nearby. Insects near or within a stream are directly affected by any chemical, physical, or biological changes in the water. An insect’s presence can be influenced by varying amounts of environmental and anthropogenic factors. As a result, this means that bats are very dependent and sensitive to the management practices of the park. Therefore, our review paper seeks to answer the question of how does regulated water releases from hydropeaking of the Tuolumne River affect bat populations and trophic food webs? Our research hopes to conclude that sporadic hydropeaking flows will negatively impact bat populations due to the displacement and reduction of aquatic insects.
Background Information
The construction of dams are a magnificent feat of human ingenuity, allowing us the ability to store water and generate hydroelectricity power. Although they provide many benefits to society, dams also generate several environmental costs. These costs negatively affect biotic factors such as salmon reproduction and abiotic factors like temperature and flow of the river. Hydropeaking is a term used to describe the pulsating flows of a river as a result from water periodically being released by a dam. Hydropeaking affects the volume, timing, and temperature of flows. The water released comes from the bottom of the dam, which is typically colder than the average river temperature, thus changing the overall water temperature downstream (Gordon 2013). Additionally, the height of the water downstream depends on the magnitude and frequency of hydropeaking water releases. The pulsating flows as a result of hydropeaking rapidly and constantly uncover or flood riparian habitat throughout the day depending on the water height. The frequent fluctuations in water level and temperature creates a volatile environment for insect populations, who depend on riparian habitat and stable water temperatures for reproduction and spawning. Hydropeaking dramatically changes riparian habitat daily with unstable conditions which threatens insect populations that reside there.
Aquatic insects are sensitive to discharge and thermal changes caused by hydropeaking. They are susceptible to both the timing and magnitude of downstream flows, as well as the temperature of the water releases. A large discharge downstream can cause disturbance known as catastrophic drift. This occurs when a large flow release flushes away the fine detritus and also some individuals living in it and leaves the remaining individuals exposed (Bruno 2013). Additionally, the temperature difference between water released from the bottom of the dam mixing with water already downstream changes the overall stream temperature, thus causing a behavioral drift. Behavioral drift occurs in response to the abrupt change in temperature causing aquatic insect species to relocate to areas where the temperatures are more favorable (Bruno 2013). Therefore, monitoring water levels by measuring gauge height and recording temperature can help predict whether a catastrophic drift or behavioral drift will be experienced by the aquatic insects located downstream.
Many studies that monitor the feeding and roosting sites of bats show that water plays a critical role in the local ecosystem for bats (Miller 2003, Seidman 2001). Recorded in the National Park Service’s survey in Poopenaut Valley, the species of bats found within the park forage directly over or adjacent to the water (Stock 2015). Therefore, the fluctuating water height, temperature, and turbidity affect bats even when not observed immediately following hydrologic changes.
Monitoring Methods
The bat’s elusive nature make them extremely difficult to monitor but not impossible. The first step is to know where to look for them. In Yosemite, bats are commonly found tucked in rock crevices or tree cavities. However, bats are known to abandon an area when the environmental pressures overwhelm them or make the habitat no longer suitable – whether it is a lack of roosting sites or a depletion in their food sources (Seidman 2001).
A common method used to monitor bats is mist netting, a process where a thin net is set up in an area where bats are known to occur. The bats fly into the net where they are collected and identified and when finished are then released. Mist netting can be dangerous for bats, because there is a chance they can be physically harmed in the process. Mist netting also requires open spaces for the bats to forage in, which limits researchers to only a few specific species (Seidman 2001). Telemetry is an alternative monitoring method, but this approach still involves mist netting and is both time and labor intensive.
One highly effective but labor intensive method is daytime roost mapping. Bats are nocturnal, making them easier to spot and count during the day when they are inactive. Roost mapping can be an effective monitoring technique, but bats do not always use the same roosts (Brigham 1997). Recounting the same bat in different roosts can pose a potential monitoring error.
Recent technology has allowed for the acoustic detection of bats. Recordings can be run through a specialized software program which identifies the bat to its species. The one downfall to this method is that the detector is not able to differentiate individual calls from different bats or repeated calls from the same bat. This makes it difficult to determine abundance of a single species, but allows for species richness to be calculated. Between roost mapping and acoustic detection, no technique is better than the other, but when deployed together it can be effective in determining bat species abundance and species richness within an area.
Analysis
By examining hydrographs with respect to the changing height of the river, we can predict whether or not insects will experience catastrophic or behavioral drift. Following the change in insect distribution, it is expected to see a correlation between bat distributions that will also change in tandem with insects.
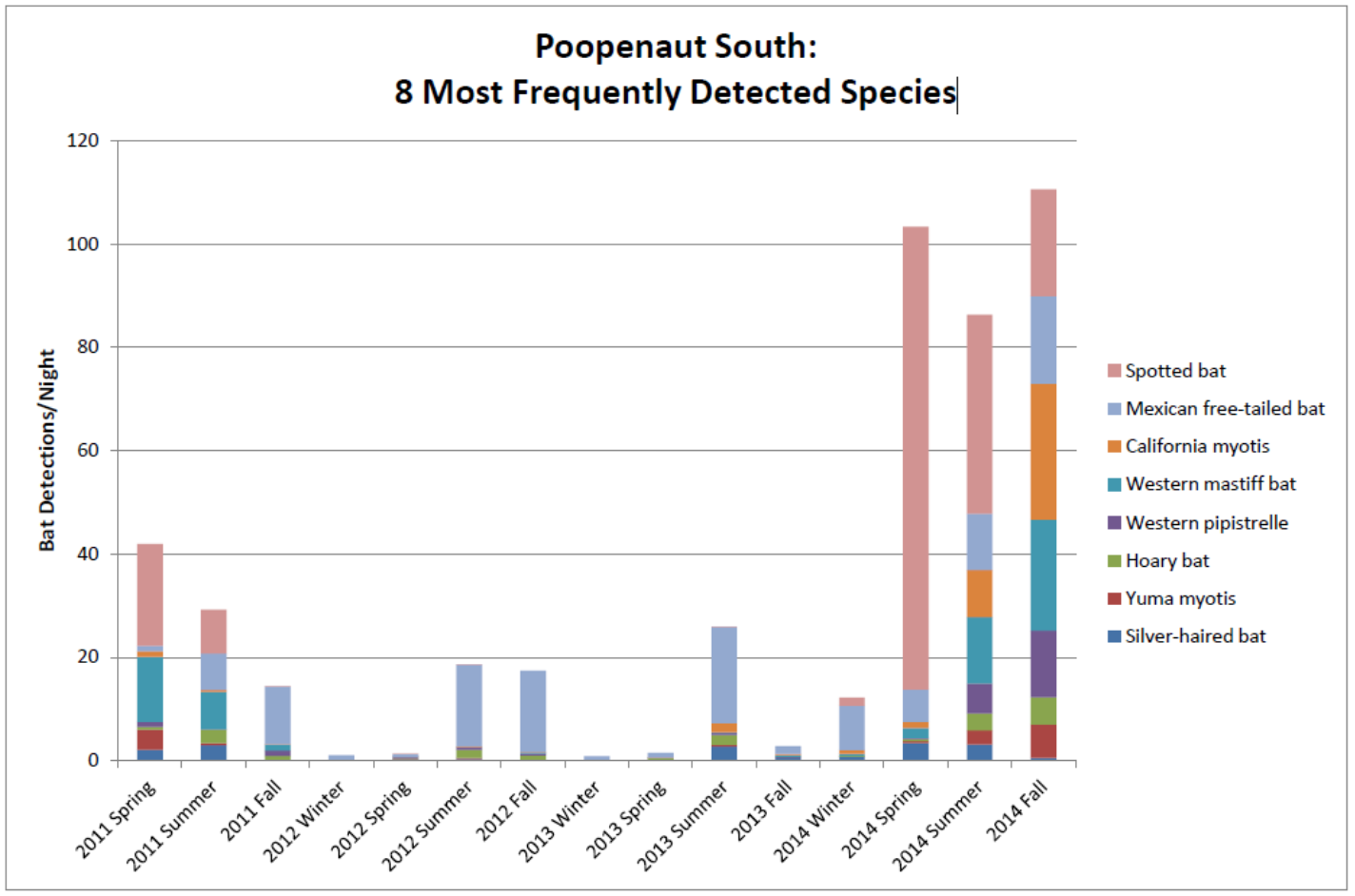
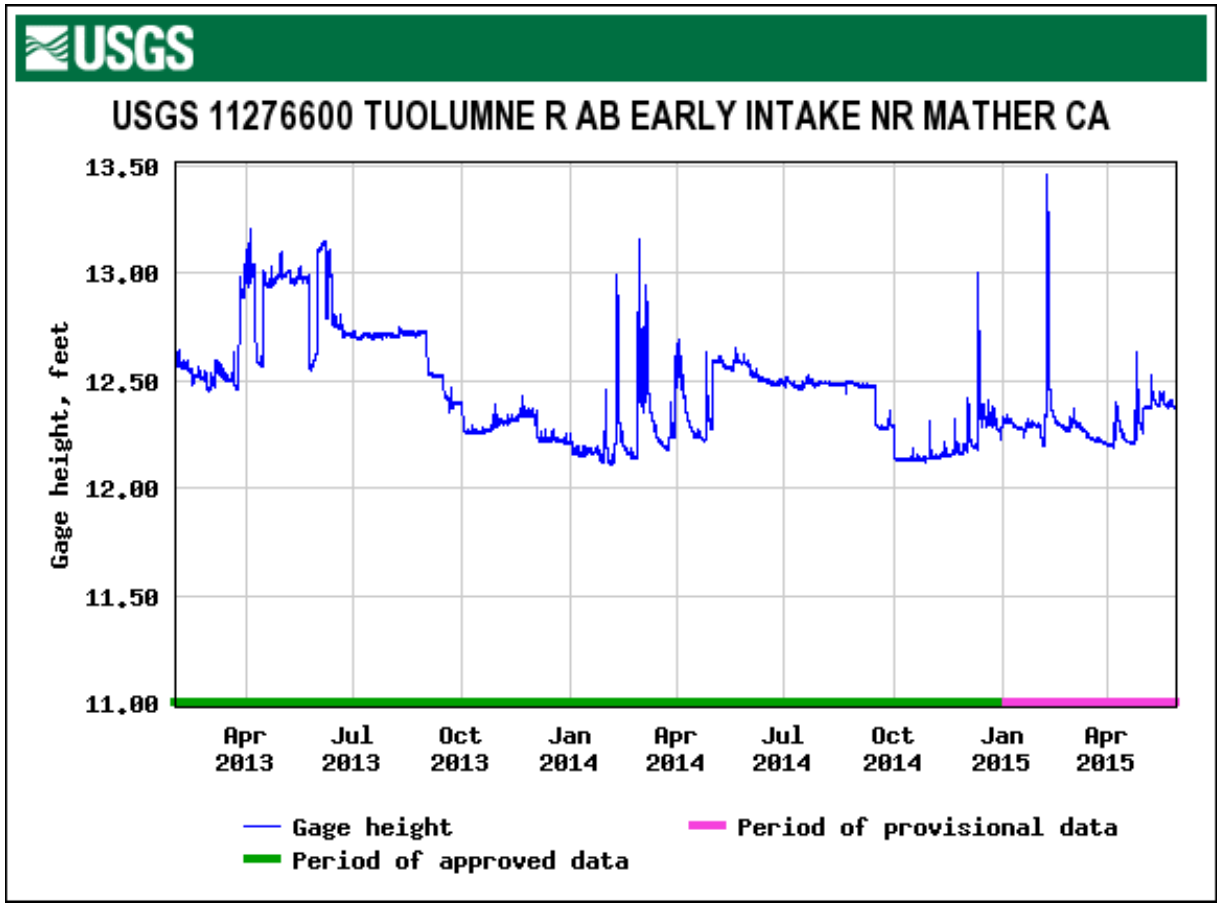
Figure 1 shows the abundance of the 8 most frequently detected species in the South Poopenaut Valley. Figure 2 is a graph generated from USGS data which reveal gauge height versus time above Tuolumne River Early intake near Mather, CA. There are two periods of relative gauge height stability around July 2013 and 2014. Summer 2013 and 2014 also happened to have an abundance of bat detections in the Poopenaut Valley. So far, this analysis is congruent with the prediction that insects are catastrophically affected by dynamic water levels. In October 2013, there is a dramatic drop in water height, and in Fall 2013, there was a significant reduction in the amount of bat sightings in the area. This rapid change could be attributed to decreased water levels, which exposed insect larvae to the elements and other predatory species. Therefore, these insects were unable to reproduce and there was a shortage of food for bats. Moreover, looking at the drastic water changes between January 2014 and April 2014, there was an astonishing amount of detections of bat species in Spring 2014. This abundance could be another source of evidence to the effects of catastrophic drift. The rapid fluctuation and large flow of discharge upstream could have caused the water to flush away insect population downstream. As a result, the bats would be able to feast on the new arrival of washed away and exposed prey. These analytical conclusions show that there could be a correlation between hydropeaking releases and bat sightings based on the theory of impacts on aquatic insects. However, these conclusions may exhibit several sources of error due to monitoring technical difficulties and an unproven correlated relationship assumption between hydropeaking flows and bats.
Conclusion
Based on our limited research, we conclude that fluctuating water levels coupled with changing water conditions affect the lowest level of the trophic cascade, insects. The effects on insects can be categorized by the response seen in the distribution of invertebrates along the river. Insects respond to the discharge level or the temperature of the flow from dam releases. Hydropeaking flow magnitude and timing can cause catastrophic drift - which displaces the insects. A sudden change in stream temperature as a result of mixing can trigger behavioral drift. The change in temperature causes insects to relocate to areas with a suitable habitat and better temperature conditions. These changes not only alter distribution and location of insects, but also influence the presence or absence of bats. By managing hydropeaking releases, we can ensure a stable insect population and contribute to the thriving bat population in Yosemite National Park.