From the data that was collected, it is difficult to say if the qualitative model outlined by Milner and Petts (1994) would apply to the Copper River and its tributaries. The large scale, of the main river channels, made thorough sampling of the entire cross section of the channel impossible and only channel margins were sampled in these areas. Samples taken in the small tributaries are likely to be more representative of the populations present, due to more thorough sampling of a larger proportion of the channel cross section. In general the data from the smaller tributaries fit the Milner and Petts model. Non-glacial tributaries showed greater abundance and diversity than the glacial tributaries, particularly when temperatures were similar between the two streams. True distributions in the main channels of the Kennicott, Nizina, Chitina and Copper Rivers proved to be difficult due to their large scale and high discharges. This probably resulted in the exceptions to the models that were observed in the main channel habitats. Distributions may fit the model more closely later in the season when discharges drop.
The distribution of macroinvertebrates in the main channels of the Copper River and its tributaries showed little in the way of patterns. In the strictest interpretation of the data, there is a weak adherence to the model, but so many pieces of the picture are missing, that anything could be interpreted from the data. There were no invertebrates collected at the first site on the Kennicott River where the temperature was .5°C, the critical temperature range from 2-4°C was passed over in one jump, at the confluence with the Nazina River, creating a large hole in the data for the main channel habitats. Beyond the confluence of the Kennicott and Nazina Rivers, the temperature increased and fluctuated between 2.8°C and 11°C. This fluctuation in temperature did not show corresponding changes in the families of invertebrates present. Chloroperlidae and Beatidae were more often found in the main channels than Chironomidae, despite the fact that the opposite was expected. It is possible that this exception was due to differences in resources available in the main channels, but sampling error would be easier to believe.
The Glacial Tributaries revealed a somewhat nicer picture. The Goodwin Glacier Distributary was the coldest of the glacial tributaries sampled. If 3.2°C was the upper limit of this streams temperature range, then Chironomidae and Simuliidae would be the only families expected under such conditions. A lack of fine particulate organic matter would explain the absence of the Simuliidae, leaving only the Chironomidae that were found there. At slightly higher temperatures, the Beatidae and Chloroperlidae would begin to show up and that is in fact what was observed in the Childs Glacier Distributary. Overall the glacial tributaries seemed to fit the paradigm because Chironomidae were the dominant family found in the samples that were taken. With the limited number of data points that were collected, little more can be said about these distributions.
The non – glacial tributaries echoed the results of their glacial counterparts. The ratios of families present in the samples generally matched the predictions made by the Milner and Petts (1994) model. Chironomidae were in the majority with Beatidae and Chloroperlidae second and, select Trichopterans third in small numbers. An increased level of diversity was observed in the non – glacial tributaries, also an expected outcome, due to reduced levels of disturbace, turbidity and increased temperatures.
When consideration is given to the difficulty involved with sampling the main channels, the differences in the results of the data are understandable. However, it is possible that the main channel habitats just don’t fit the paradigm described by Milner and Petts (1994) and the differences in scale are more significant than previously thought. The extreme levels of sediment load carried by the Copper River and its tributaries are likely to be one of the main factors influencing the discrepancies that were observed. Additionally, the presence of adult forms, where no larval forms could be found, indicates error in sampling techniques. This idea is supported by the good data and results that were collected for the smaller tributaries.
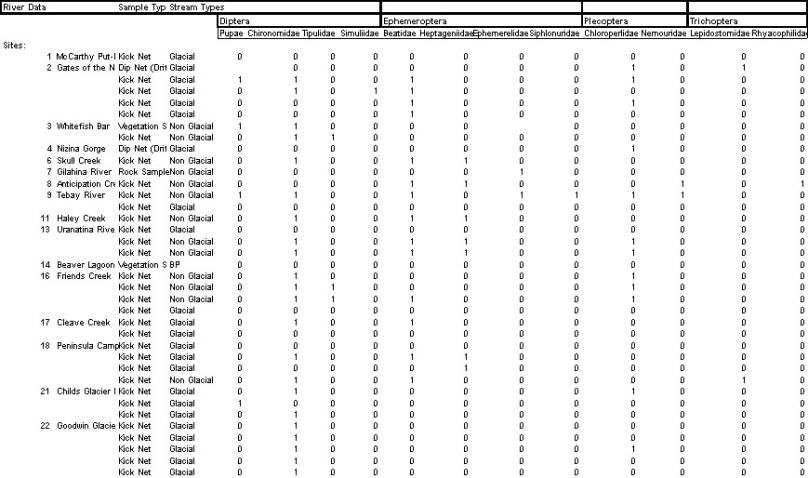
Site 1: McCarthy
The Kennicott River was at or approaching it’s annual peak at the time of sampling. This would imply the highest levels of disturbance and lowest temperatures for the year. The lack of insects in the sample would apparently be related to the water temperature, which was .5°C at the time the sample was taken. This extremely low temperature would completely exclude all insects from living in this habitat, at this point of the year.
Site 2: Gates of the Nazina
This site revealed one of the interesting exceptions to what I expected to see. Samples taken from the main channel of the Nazina yielded only the stonefly, Chloroperlidae. It is not clear if the individuals sampled were originating from the main channel, because they did not turn up in kick samples, only in dip nets at the waters edge. The temperature below the confluence with the Kennicott River was 4°C and the disturbance was assumed to be high, therefore, samples should have been dominated by Chironomidae. The fact that Chloroperlidae are predators only adds confusion to these results, as no prey of any sort were found in the kick net samples either. The possibility of movement from the side channel, in the drift, was weakened by the fact that samples taken above the side channel were similar to those taken below it. This is compounded by the fact that no Plecoptera were found in the clear side channel.
The hyporheic flow channel samples showed an expected increase in diversity, but still less than I would have expected to find. At 14.7°C and very low disturbance, I would have expected to find a very rich diversity of insects in this location. This site was rich in nutrients as demonstrated by the high levels of algae and periphyton growing in the pools. Under these conditions I would have expected to find representatives of all of the EPT, yet Plecoptera and Trichoptera were missing.
Site 3: Whitefish Bar
This site was a well protected, hyporheic channel that was not subject to a high frequency of disturbance. The considerable sizes of the willows growing on all parts of the bar and around the stream channel were indication of this low level of disturbance. The water in the hyporheic channel was cooler than the water in the hyporheic channel at Gates of the Nazina, yet they were still warm enough for all of the invertebrate orders that we might see in a glacial stream. Chironomidae were the most numerous group in the samples that were taken with Tipulidae second. The presence of Tipulidae indicates that this small channel is quite stable with low levels of disturbance on an annual scale. I would expect that more thorough sampling in the riffles would have turned up members of the EPT, the stoneflies in particular, as they fit a similar trophic niche as the Tipulidae.
The vegetation sweep only contained Chironomidae, which coincides with the findings of the kick net samples. Chironomidae are the more mobile of the two families found in the riffles and prefer grazing the algae that was growing on the submerged vegetation.
Site 4: Nizina Gorge
The presence of Chloroperlidae in the flotsam at this location points to movement in the drift. The river in this location was focused into one deep channel with considerable gradient and very fast water. I was not able to take a kick net sample due to the depth and velocity of the water, and individual examination of rocks did not yield any specimens. This evidence points to the drift as the source of these individuals in this location.
Site 6 and 7: Skull Creek and the Gilahina River
Skull Creek is a non-glacial tributary with high gradient and clear, relatively warm water. This stream was the first to have Heptageniidae. The non-glacial nature of this stream makes it a prime habitat for Heptageniidae. The site sampled was approximately .25 m deep with fast water and medium sized cobbles. This is exactly where Heptageniidae would be expected to be the dominant grazer, over Chironomidae. Beatidae were also present but in smaller numbers.
The Gilahina River was similar to Skull Creek, yet the invertebrates found there were of a different family of Ephemeroptera. Temperature, water quality and habitat were quite similar between these two streams. One difference was the substrate. The Gilahina was much more armored than Skull Creek, having medium to small cobble that was very difficult to displace. This could have been the difference in habitat that was responsible for the difference in taxa that were found. By turning over rocks, I found only Ephemerelidae, a large grazing Ephemeropteran. These were no doubt grazing on the limited amounts of periphyton that was growing on the rocks.
Site 8: Anticipation Creek
Anticipation Creek is another small non-glacial tributary that drains into the Chitina River. At 7°C the temperature was not significantly different relative to the other non-glacial tributaries that were sampled. The lack of a braided delta at its confluence with the Chitina was similar to Skull, Tebay, and Haley Creeks. The gradient was quite steep and continuous for the entire stretch that was observed. A unique character of the site sampled was the dense canopy over the stream, if this was consistent for the length of the stream, it would have negative impacts on algal growth. The combination of lowered algal growth and high leaf deposition would create excellent habitat for Nemouridae, the Plecopterans capable of digesting cellulose. This was in fact what I found. The absence of Chironomidae may not have been related to low levels of algae production, because the Heptageniids were doing just fine. But, this was the first location that I found Nemouridae, probably due to the unique combination of water quality and high levels of CPOM.
Site 9: Tebay River
This site was unusually rich in diversity among all of the sites sampled. The Tebay River is a non-glacial river with low turbidity and of average temperature among the non-glacial tributaries sampled. It was also one of the largest tributaries sampled. The substrate of the site sampled was medium sized cobble with little sand in between, and high levels of CPOM, presumably caught in the interstitial spaces of the cobble. With six different families in one sample, this would appear to be a relatively rich system. Families present included Chironomidae, Beatidae, Ephemerelidae, Siphlonuridae, Chloroperlidae, and Nemouridae. The high diversity among the Ephemeroptera and Plecoptera indicate good water quality and high levels of dissolved oxygen. A Kick net sample taken in the main channel of the Chitina River above the confluence with the Tebay River did not produce any specimens.
Site 11: Haley Creek
The confluence of Haley Creek with the Copper River was similar to that of Anticipation Creek, the main channel did not have a braid plain at the confluence. The sample was taken just downstream of a waterfall that is most likely a barrier to migration for most fish. The substrate in this location was large cobble and actual kicking for agitation was quite difficult. Chironomidae, Beatidae and Heptageniidae were all collected in the Kick Net sample and due to the inefficiency of the sample taken it is likely that there were more species present. It is interesting to note that gut content analysis of the Greyling sampled at this site revealed that the fish were not feeding on the aquatic insects present in the stream. They were instead feeding heavily of terrestrial forms that were present in the drift, largely in the foam at the surface. Even the unskilled casting of a complete novice was able to entice one of these voracious predators into striking at a fake. The high concentration of food on the surface must have been an unusual occurrence to cause such aggressive feeding.
Site 13: Uranatina Creek
This site had a uniquely large braid plain at its confluence with the Copper River. Therefore, while this tributary is medium to large in size, the habitats sampled were on a rather small scale, taken individually. The composition of taxa at this site was very much like that of the other non-glacial tributaries, dominated by Beatidae and then Chironomidae and Heptageniidae. These three seemed to be the most common among all of the tributaries. Chloroperlidae was also present in the samples. A sample taken in the main channel of the Copper River above the confluence did not yield any macroinvertebrates for identification.
Site 16: Friends Creek
This was a very small, short and consequently steep non – glacial tributary to the Copper River. 6.6°C made it about average temperature for the non – glacial tributaries. The taxa found here included Chironomidae and Chloroperlidae. The exceptional taxon was Tipulidae. They were found in small channels with large and medium gravel and swift water with standing waves. This was not a unique type of habitat among the areas sampled yet the presence of Tipulidae is quite unique. However, the temperature and water qualities of this site and the Whitefish Bar were similar which is consistent evidence for the conditions that Tipulidae need to survive.
Site 17: Cleave Creek
This was the first of the glacial tributaries that were sampled. The confluence with the Copper River was very different from those of the other tributaries. The evidence of disturbance was greater, having a larger flood plain and less vegetation growing between the braids gave the impression that this channel saw large flood events frequently. The species represented in the kick samples matched this impression. At 3.6°C, Chironomidae and Beatidae are the families most likely to be present, of those observed in this system. The Milner and Petts (1994) model would predict that there would be Simuliidae in this type of stream, and Chloroperlidae and Nemouridae as well.
Site 18: Peninsula Camp
This was a large and complex site to sample. The main channel samples on the Copper River were consistent with samples taken all along the length of the river, no macroinvertebrates. The samples taken in the smallest of the main channel braids were the first main channel habitats to yield invertebrates since the Nazina Gorge. The very small channel that was connected to a clear tributary channel yielded Chironomidae, Beatidae, and Heptageniidae. While this was technically a main channel habitat, it was more similar in its physical features to that of a tributary braid, with its medium gravel, narrow channel and slower current. While this channel may not last for more than this one season, for the time being it is stable habitat for these macroinvertebrates. It is most likely that these individuals moved into this area from downstream in the clear tributary.
The clear tributary was literally the water of the Copper River that had been filtered via hyporheic flow and warmed in a Beaver Pond at the upper end of the peninsula. The stability and warmer temperatures of this habitat are reflected in the presence of the Trichopteran, Lepidostomidae. These along with Chironomidae and Beatidae were found in the sample taken at this site.
Site 21: Childs Glacier Distributary
This site was the closest of any of the sites to the originating glacier. The area sampled was the edge of a riffle in the largest channel of the tributary. The temperature was 5°C in the distributary and the substrate was mostly medium cobbles. The turbidity was much more similar to those reported by Milner and Petts in their work, at 45.8 NTU. This site should have best matched the model, and did in ways. The sample taken only yielded Chironomidae and Chloroperlidae, but this was one sample during the peak of the season. It is possible that further up the channel in areas where the water was slower, the diversity and abundance could have been higher, in accordance with the temperature. Even still, the Families represented in this sample are in accordance with the predictions made by the model. The absence of other families does not prove anything about the accuracy or validity of the model.
Site 22: Goodwin Glacier Distributary
This was the coldest of the glacial tributaries sampled, at 3.2°C. The water quality was similar to that of the Childs Glacier Distributary. The gradient was low and there were many braids and shallow riffles were abundant. The sites sampled all had small to medium gravel and plenty of algae. This was optimal conditions for Chironomidae to dominate, and that is exactly what samples returned. Closer analysis would have likely shown that the Chironomids present were from the sub – families Diamesinae and Orthocladinae, according to the model by Milner and Petts (1994). No other taxa were found in four samples.