- INTRODUCTION
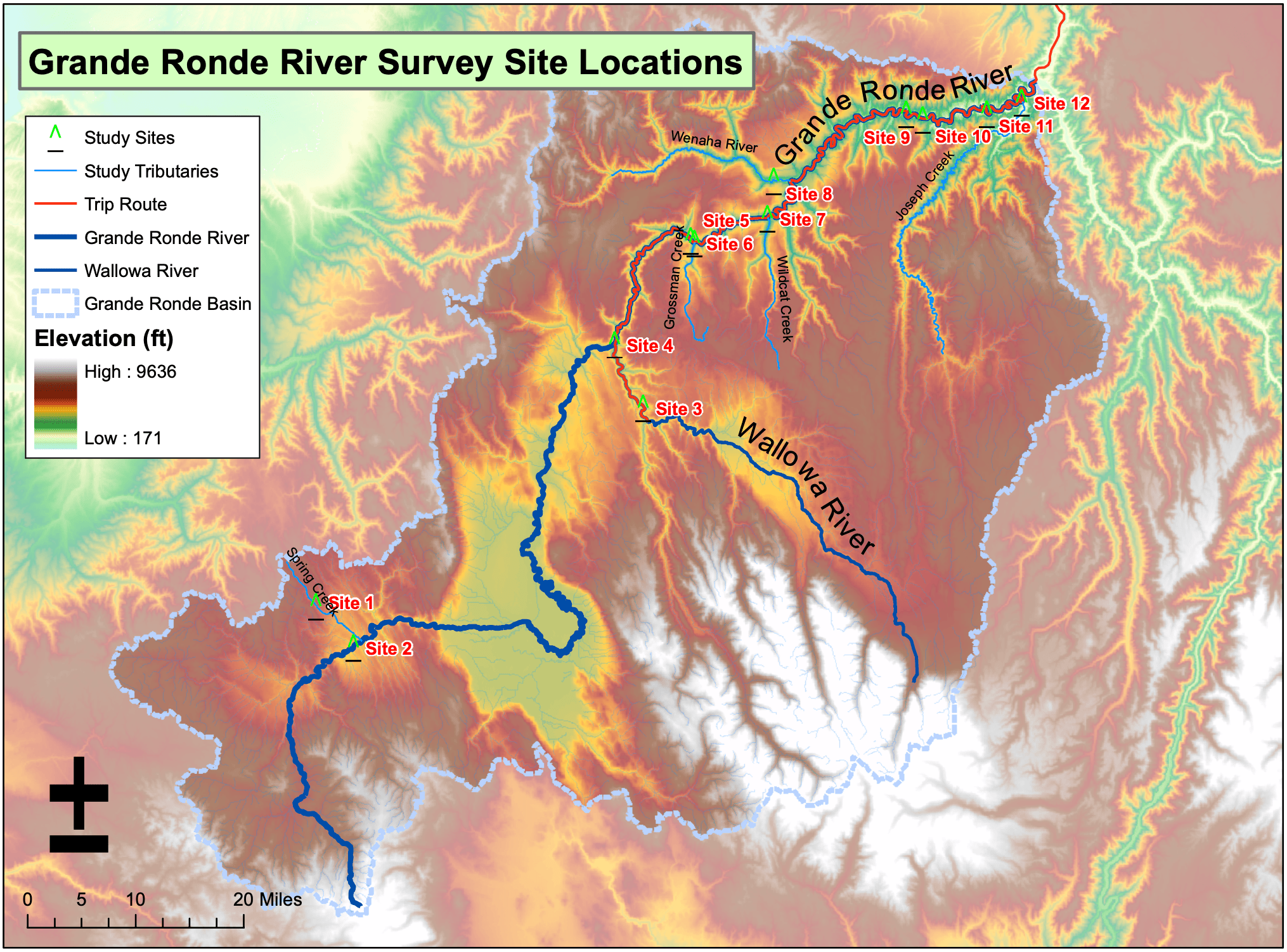
Vannote et al.'s (1980) River Continuum Concept (RCC) describes the biological community structure and function of a river from its headwaters to mouth as a continuous, longitudinal gradient controlled by a longitudinal gradient of physical features. We conducted simple surveys (June 19-June 26, 2007) along the Grande Ronde River, in northeast Oregon/southeast Washington, USA, from headwaters to mouth (Figure 1), in order to test our hypothesis that this continuum would be observable along the Grande Ronde. It is one of the few rivers in the Pacific Northwest that remains undammed. Historical intensive logging, as well as current logging, ranching, and irrigation agriculture comprise major land uses. However, a significant stretch of the Grande Ronde is a designated Wild and Scenic River, which minimizes the observed anthropogenic impacts along our study reach. To test our hypothesis, we mapped hydrogeomorphic patches, tracked local meteorological conditions, and recorded woody vegetation and wildlife species diversity.
- EXPERIMENTAL DESIGN AND METHODOLOGY
Survey sites were chosen at different hydrogeomorphic patches, after Thorpe et al. (2006) (see Hestir 2007, this volume). General geomorphic process zones were identified using USGS 1:24,000 digital raster graphics topographic maps, and hydrogeomorphic patches were selected while in the field. Figure 1 depicts the final survey sites selected. Geomorphic process surface maps were created for each of the survey sites (Figures 2-13). Air temperature, relative humidity, barometric pressure, and wind speed were measured using a handheld weather meter. Vegetation was characterized along transects, and by conducting woody species inventories. Canopy cover and elevation were also recorded along the transects. Transect locations can be found on the accompanying site maps. Additionally, woody species inventories were taken for each survey site. Bird diversity was determined by conducting timed point counts as well as noting other bird species observed at surveyed sites, campsites, and traveling along the river. Mammalian diversity was determined through observations of species based on tracks, scat, and sightings. Diversity of amphibians and reptiles was determined with timed searches in which rocks and logs were overturned and other sightings at surveyed sites.
- RESULTS
The geomorphic process surface maps were used to develop a qualitative description of the change in physical features along the river. Although the geology of the basin does not significantly change along the river, geomorphically the river does demonstrate a downstream gradient. As we moved from headwaters (Figures 2-4) to mouth (the confluence with the Snake River) (Figures 10-13), the antecedent river features became more pronounced as the river increased in magnitude. The canyon walls became more constricted, and the occurrence of bedrock in the active channel increased in the downstream direction. However, the observed gradient was punctuated by small scale discontinuities created by hydrologic processes and local scale geomorphic processes such as changes in slope, channel constriction ratios, and tributary-mainstem interactions. In the middle reaches of the river, tributaries (i.e. Grossman Creek, figure 7; Wildcat Creek, figure 8; and the Wenaha River, figure 9) were influential factors in geomorphic processes and surfaces in the mainstem channel (Grande Ronde). Tributary-mainstem interactions were characterized by large cobble/boulder creek-fan surfaces that tended to constrict and deflect primary flows in the Grande Ronde. Channel gradient remained relatively consistent throughout the upper and middle reaches of the study stretch. In the lower reaches, gradient decreased noticeably, with the exception of one significant slope increase formed by bedrock constriction of the channel ("The Narrows" rapid).
Climatic conditions form part of the physical template of a riverine ecosystem (sensu Vannote et al. 1980). Observations along the Grande Ronde (Figure 14) show that temperature did not follow a predictable trend in the downstream direction, but rather changed due to general variability. On the other hand, humidity generally decreased in the downstream direction, with some stochastic variability, consistent with casually observed changes in non-riparian vegetation. There was a notable correlation between recorded maximum daily air temperature and maximum daily stream temperature, as noted by Fissekis et al. (2007, this volume). Wind speeds were generally below 5 mph, and barometric pressure increased steadily from 1005 mb in the headwaters to 1091 mb at the mouth.
Longitudinal trends in the woody vegetation composition and richness are apparent. Species richness clearly peaks in the middle reaches, consistent with the intermediate disturbance hypothesis (see Hayden, 2007, this volume) (Figure 15). We observed a marked downstream gradient in vegetation architecture, from coniferous forests in the headwater areas, to woodland, savannah, and finally bunch grass/steppe at the mouth. Land use impacts were evident above and below the Wild and Scenic section. This included logging above, and grazing below the designated reach. Cattle grazing along the river bank significantly reduced the presence of riparian vegetation such as willow, and an increase in invasive species was observed. An exploratory survey of large woody debris frequency along the river gradient followed a similar pattern as the woody vegetation gradient. See Lawson flog (2007, this volume) for a detailed account of this investigation.
Vegetation presence, topographic profile and canopy cover were all highly dependent on micro-patch location, but longitudinal trends in variation were observed between those locations. Figure 16 depicts the distribution of riparian vegetation along transects. Our results demonstrate that vegetation types follow trends in elevation above the river. For example willow (Sa/ix spp.)were generally restricted to in range to less than one meter above the river, alder (A/n us spp.)and Dogwood (Corn us spp)were found at intermediate elevations, and the range of snowberry (a deciduous shrub, Symphoricarpos spp) and pine trees (Pinus spp.)extended far above the river. Generally, canopy cover decreases in the downstream direction.
The diversity of riparian vegetation along the Grande Ronde River is a useful proxy for habitat diversity and thus wildlife diversity and richness. Wildlife species richness, defined as the number of species for a given area, did not follow a general longitudinal gradient for the terrestrial species we studied. Amphibian, reptile, and mammalian richness did not follow a longitudinal gradient but rather remained relatively constant throughout the surveyed sites (Figure 16). Bird diversity was highest in the middle reaches, primarily in concurrence with the presence of higher proportion of edge habitat, such as the presence of riparian zones alongside a flood terrace grassland. Both the number of functional groups (see Appendix A) as well as the proportion of each group represented changed in the downstream direction (Figure 17). The middle reaches, for example, had the greatest number of functional groups and some of the dominant functional groups in the headwaters and lower reaches were smaller in proportion to overall diversity.
This pattern is inconsistent with the predictions made by the RCC; it does not demonstrate a continuous downstream gradient. Species diversity was highest in areas within the middle reaches such as Wildcat Creek (Figure 8) which are composed of many different habitats including forested areas, open grassy areas, woody deciduous riparian vegetation and edge habitats created by both anthropogenic and natural processes. These edge habitats may be indicative of a punctuated or discontinuous downstream gradient.
- CONCLUSIONS
We conducted geomorphic surface mapping, weather and wildlife observations, and vegetation surveys, in order to test whether terrestrial facets of river systems demonstrate a continuous gradient from headwaters to mouth. The results of these surveys lead us to conclude that:
- There was an observable downstream gradient of geomorphic features from headwaters to mouth along the Grande Ronde River. However, this gradient is scale dependent; at the scale of the hydrogeomorphic patch, this gradient disappears. Furthermore, this gradient is best characterized as discontinuous, where tributary-mainstem interactions create hydrogeomorphic punctuations along the gradient.
- The downstream gradient of geomorphic features did not result in a continuous downstream gradient of ecological structure and function along the river. The highest levels of biocomplexity were observed at the middle reaches of the river, forming a bell-shaped distribution as predicted by the Intermediate Disturbance Hypothesis (see Hayden, 2007, this volume).
Figure Captions
Figure 1: Overview map of the Grande Ronde River Basin with survey sites. Figures 2-13: Geomorphic process surface maps of each survey site.
Figure 14: Recorded maximum daily temperatures and average streamside relative humidity
Figure 15: Characterization of Riparian Vegetation. Transect tape was laid out orthogonal to the riverbank. Elevation was measured using a laser range finder; canopy cover was measured with a densitometer; and vegetation with branches found within three meters of the transect tape were included in vegetation ranges.
Figure 16: Species richness at surveyed sites.
Figure 17: Bird functional groups in the upper, middle, and lower reaches of the Grande Ronde River. Figure 18: Total number of bird species observed during travel downstream in between survey sites.
Table 1: Woody species presence at survey sites. Woody species inventories were taken within the boundaries of the site maps. Most plants were identified down to the genus or species level, while some species—listed as unknown—were not identified. Absence of a species within the boundaries of a site map boundaries does not indicate absence of that species within the river reach. Notably, many species that were not present at site nine were present just outside the boundaries of the site map.
Table 2: Observed wildlife species in sample sites
Appendix A: Functional groups of birds observed along the Grande Ronde River