I. INTRODUCTION
The River Continuum Concept (RCC) describes a downstream gradient of organisms that follows a downstream gradient of physical characteristics. Tributaries may not share the same physical characteristics as the mainstem, creating network nodes where physical and biological anomalies disrupt the longitudinal patterns found along the mainstem river. This paper examines the aquatic environment of the Grande Ronde River to determine whether tributaries have a spike or a step effect on downstream trends in physical habitat characteristics and aquatic organisms within the mainstem. To examine changes in these trends along the mainstem river due to tributary confluences, we measured discharge, obtained water quality measurements, and sampled subs trate, invertebrates, and fish, above, below and at tributary confluences (Table 1) over the length of the river sampled.
II. EXPERIMENTAL DESIGN AND METHODOLOGY
Discharge from the tributaries, and where possible, the mainstem Grande Ronde River, was determined using standard velocity-area methodologies, with velocity measurements obtained with a Marsh-McBirney Flo-Mate velocity meter. Near-bed, instream water temperatures were obtained at each sampling site using a YSI Multimeter hydrolab, with additional water temperature data collected continuously at 30-minute sampling intervals using a thermistor attached to a raft. Channel substrate was sampled using the Wolman Pebble count method along transects above, below, and at tributary confluences along with tributaries and other field sites. Invertebrates were collected with a Surber sampler and identified to family. Fish distributions were observed by snorkel and hook-and-line sampling methods. Snorkeling surveys were conducted only at identified sampling sites, while hook-and-line surveys were carried out by individual anglers fishing both from the rafts and from the shore at our sampling sites. Sampling began at the headwaters of the Grande Ronde at Spring Creek (confluence located at river mile 173.4) and ended at the confluence of the Grande Ronde with the Snake River. A list of sampling sites and dates is presented in Table 1.
III. RESULTS
Although tributaries did contribute to the increasing discharge of the Grande Ronde, only the Wallowa River made a step change to the overall discharge in the Grande Ronde (Table 2). The Wallowa tripled the volume being transported by the mainstem river; this increased volume was maintained until the confluence of the Grande Ronde with the Snake. Generally, no other tributaries were large enough to increase the Grande Ronde's discharge by more than one percent with the exception of the Wenaha River, which increased the mainstem discharge by over 10 percent. This isolated shift implies that stream size may be related to the ability of a tributary to impact a mainstem river.
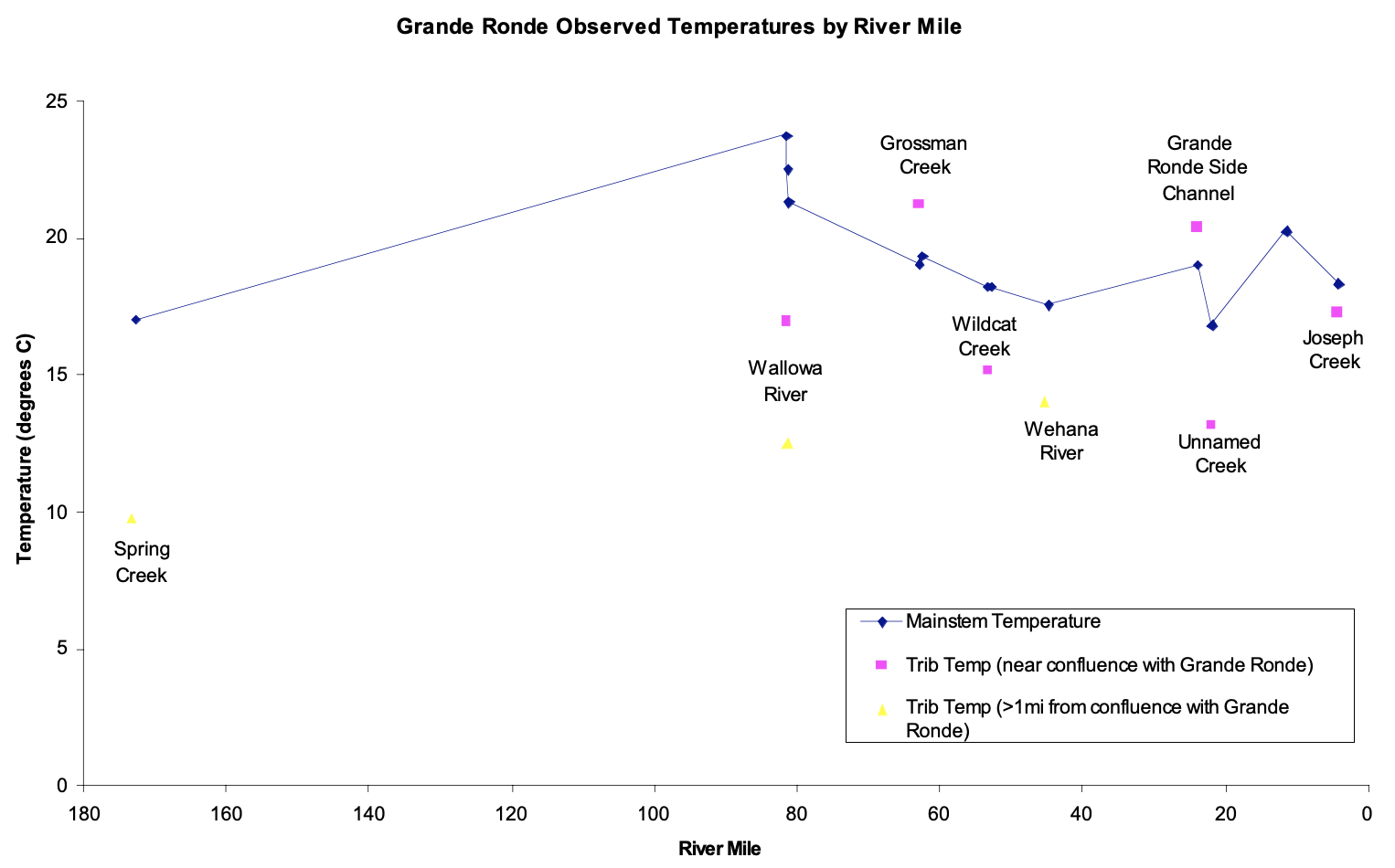
Similar to its effects on discharge, the Wallowa River also affected a step change in the temperature of the Grande Ronde River (Figure 1). The cold temperature of the Wallowa reduced the water temperature in the Grande Ronde; the Grande Ronde never recovered its original temperature levels along the rest of its longitudinal downstream gradient. Although several other tributaries were also significantly colder than the mainstem river, only the Wallowa had enough discharge to significantly lower the Grande Ronde River's temperature (Figure 1): ambient air temperature and the diurnal cycle had a greater impact on mainstem temperature shifts (Figures 2 and 3).
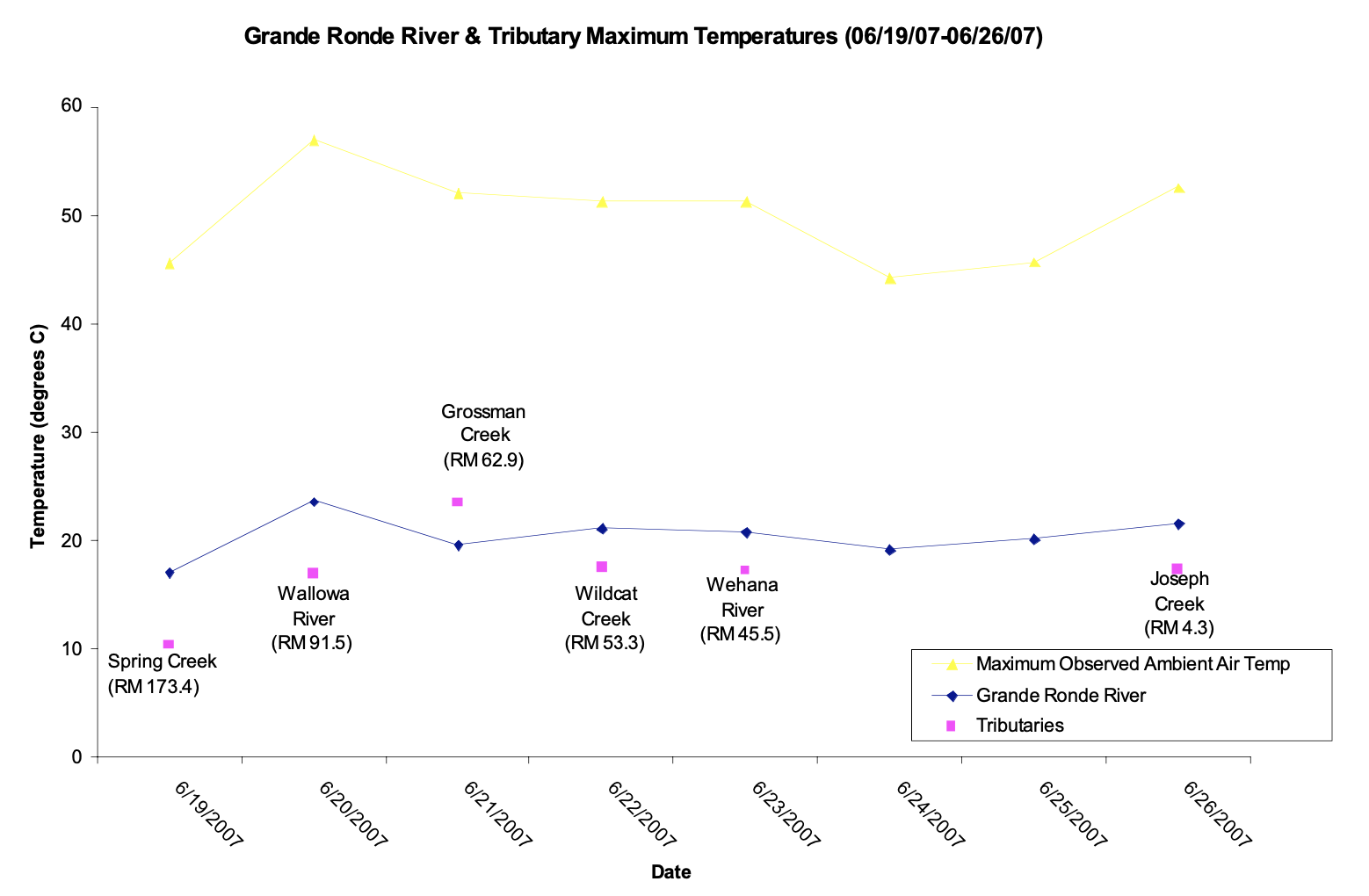
Temperature did not continually increase in the Lower Grande Ronde River because it was affected by a cold front which moved in on June 23 (Figure 2). Thus, excepting the Wallowa, tributaries had neither a step nor spike effect on the Grande Ronde River's longitudinal temperature gradient. Again, this isolated shift implies that stream size is related to a tributary's ability to disrupt a mainstem river's physical characteristics.
Conversely to the previous changes in downstream trends of physical characteristics, the results from the grain size analysis show that tributaries contributed spikes in sediment size in the downstream direction (Table 3). Sediment size disruptions at confluences are clearly seen in Figure 6. Sediment size decreased on the mainstem just below its confluences with the Wallowa River, Wildcat Creek, and Joseph Creek (Figures 7 & 10). The cause of the sediment fining is due to the deposition of fine sediment at the mouths of these tributaries and its subsequent transport into the Grande Ronde River. Localized, short-lived reduction in sediment size resulted from this deposition and ultimately negative spikes in the sediment size occurred in the downstream direction.
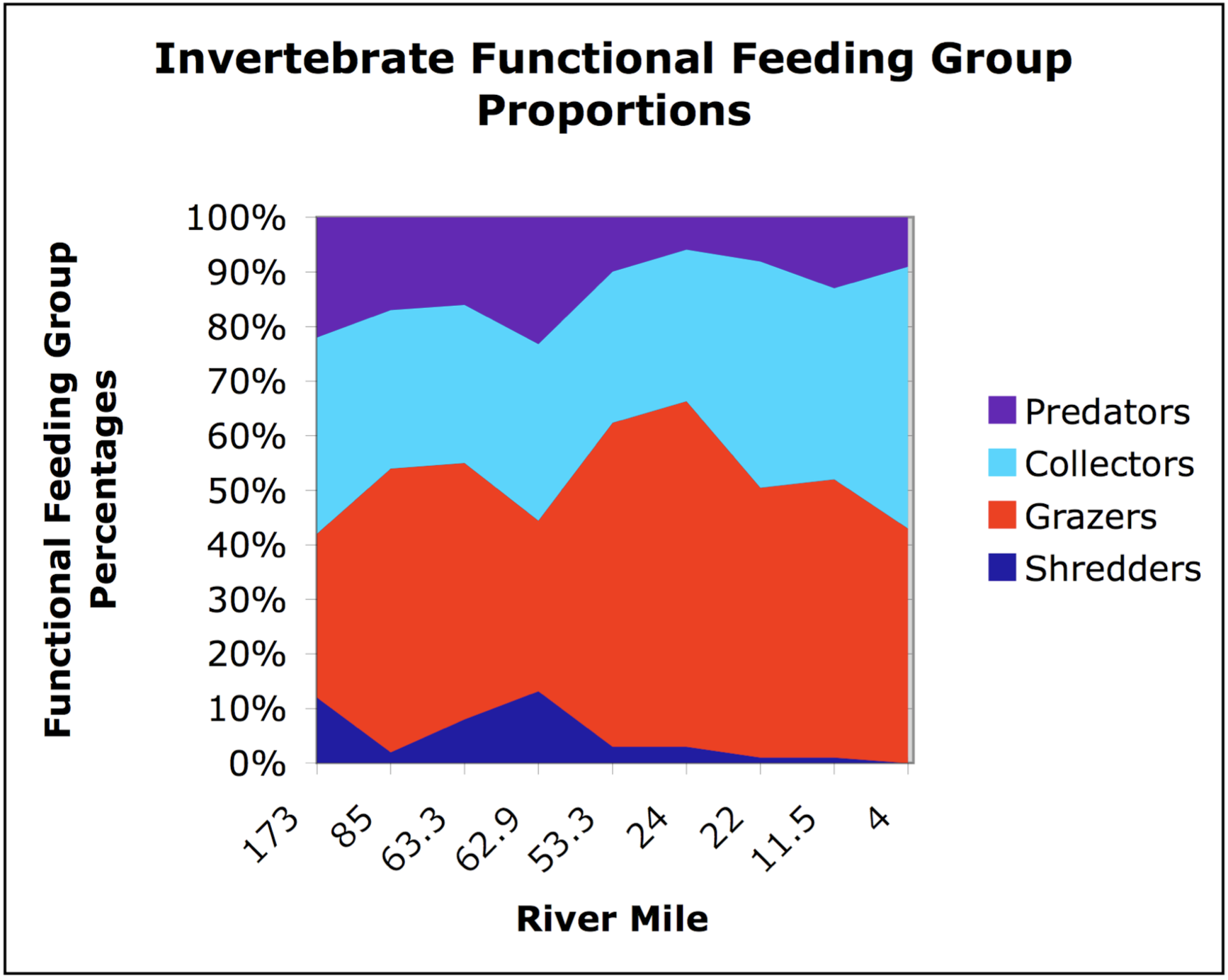
Tributaries did not seem to affect the downstream gradient of biologic characteristics. The distribution of invertebrates generally followed the trends predicted by the RCC,with most tributaries having no impact on the invertebrate fauna of the mainstem (Figures 11 and 12). For example, though pteronarcid stoneflies were present in Wildcat Creek, these insects were absent from the mainstem below the confluence (Figure 11). However, one tributary - Grossman Creek - did locally influence the Grande Ronde, as distinctive taxa(Lepidostomatidae and Perlidae) found in this creek were also present downstream in the mainstem (Figure 11).
Fish abundances and distributions changed in a downstream direction from the headwaters to the mouth of the Grande Ronde. Our results show that the upper reaches were dominated by salmonids while the lower reaches were dominated by smallmouth bass (Figure 13). Native non-salmonids occurred throughout the entire river system. Cottids, cyprinids, and catosomids were found in greater abundances in the upper reaches of the Grande Ronde; however,they were also found in the tributaries and in the downstream reaches. Our sampling techniques were not adequate to detect whatever patterns of distribution and abundance existed. Sampled sections of the Wallowa and Wenaha tributaries below the headwaters of the Grande Ronde River showed a higher abundance of salmonids compared to the mainstem of the middle and lower reaches of the Grande Ronde (Figure 14). Therefore, the Wallowa and Wenaha tributaries contributed spikes salmonid abundance in an otherwise continuous downstream decline in relative salmonid abundance. The upper reaches of the Grande Ronde mainstem showed higher abundances of salmonids than in the middle and lower reaches.
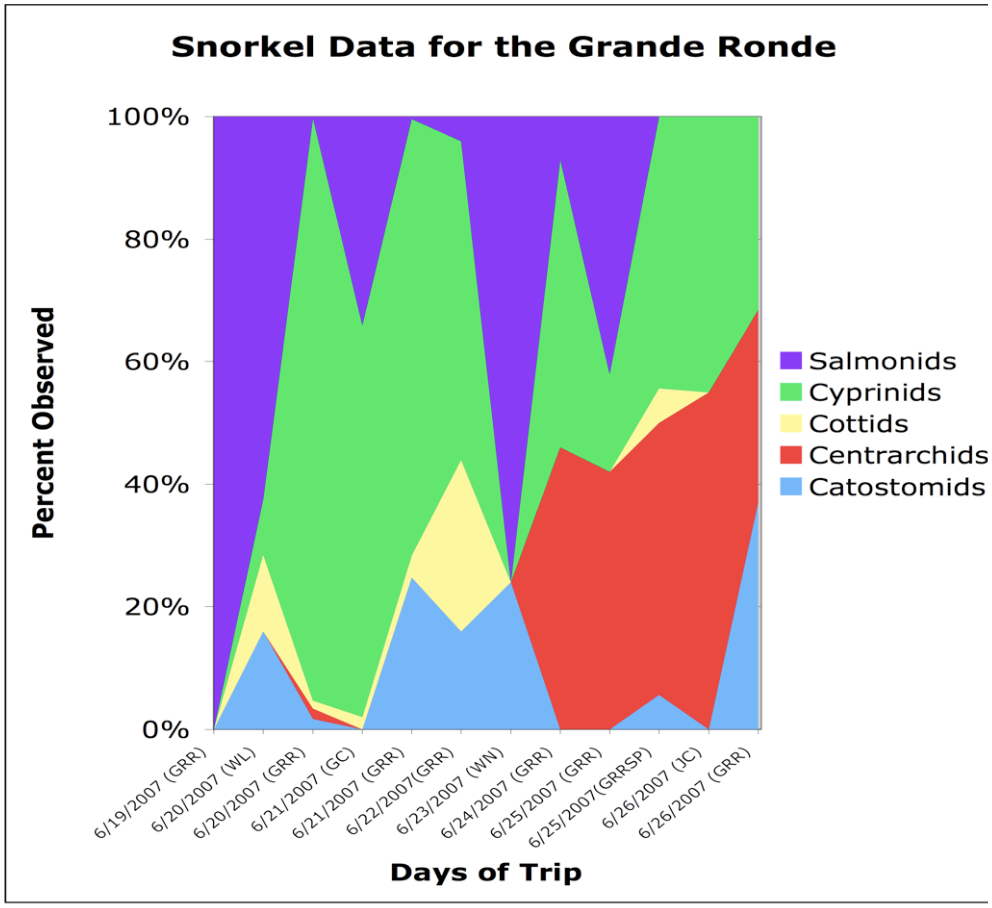
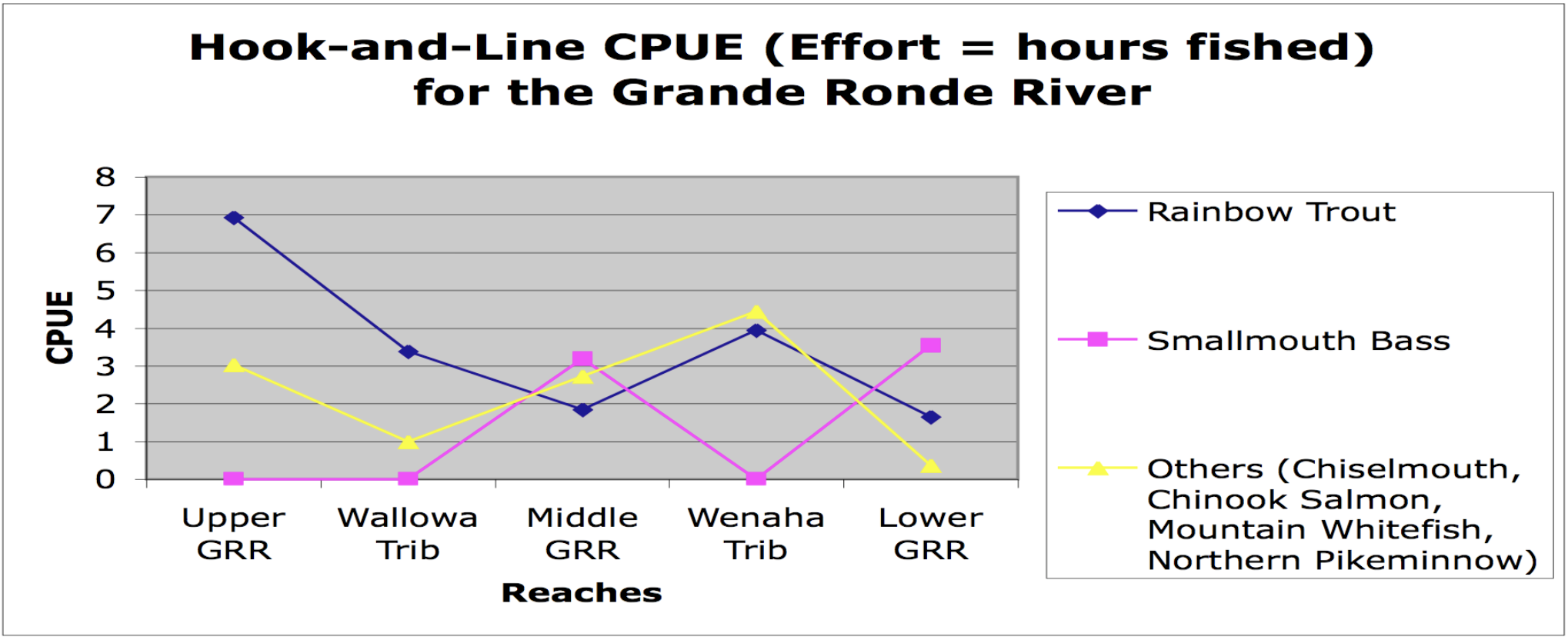
Cold-waterfishes (i.e.,rainbow trout) dominated the upper reaches, while they were gradually replaced by warm-water fishes (mainly non-native smallmouth bass Micropterus dolomieui) in the lower reaches(Figures 13 and 14). Although native cyprinids and Fish abundances and distributions changed in a downstream direction from the headwaters to the mouth of the Grande Ronde. Our results show that the upper reaches were dominated by salmonids while the lower reaches were dominated by smallmouth bass (Figure 13). Native non-salmonids occurred throughout the entire river system. Cottids, cyprinids, and catosomids were found in greater abundances in the upper reaches of the Grande Ronde; however,they were also found in the tributaries and in the downstream reaches. Our sampling techniques were not adequate to detect whatever patterns of distribution and abundance existed. Sampled sections of the Wallowa and Wenaha tributaries below the headwaters of the Grande Ronde River showed a higher abundance of salmonids compared to the mainstem of the middle and lower reaches of the Grande Ronde (Figure 14). Therefore, the Wallowa and Wenaha tributaries contributed spikes salmonid abundance in an otherwise continuous downstream decline in relative salmonid abundance. The upper reaches of the Grande Ronde mainstem showed higher abundances of salmonids than in the middle and lower reaches.
IV. CONCLUSIONS
The RCC states that lotic systems exhibit smooth, longitudinal physical gradients with concomitantly smooth, longitudinal biological gradients; implicit in this idea is that both gradients completely lack discontinuities. In general, our results show that tributaries disrupt the continuum introducing both spike and step discontinuities in mainstem gradients. For instance, the Wallowa River caused a step increase in discharge and a step decrease in the temperature of the Grande Ronde River (Table 2 and Figure 2, respectively). Additionally, the cold front of June 23 likely decreased the Lower Grande Ronde's temperatures, resulting in notemperature gradient at the basin scale (Figure 2). Substrate size also lacked a gradient for two reasons: first, the Grande Ronde's grain size was relatively constant due to the common sediment source (basalt canyon walls) for the entire river; second, tributaries caused local decreases in sediment sizes downstream of confluences (Table 3 and Figure 6). Biologically, the change in the invertebrate functional feeding group proportions followed the RCC in most respects but was disrupted at the Grossman Creek confluence (Figure 11). Fish communities also exhibited this same pattern: salmonid abundance decreased downstream, only to be interrupted by positive spikes at the confluences of the Grande Ronde with the Wenaha and Wallowa rivers (Figure 13).
Although our findings show that the aquatic features of the Grande Ronde River are not entirely consistent with the RCC, this may reflect our limited abilities to sample the river as much as problems with the RCC. The RCC should be viewed as a simple, incomplete foundation that is a useful foundation to upon which to built a more complex understanding of how lotic systems function.
APPENDIX